파이토호르몬과 식물의 2차 대사산물은 밀접한 관련이 있습니다.
옥신과 같은 호르몬은 식물의 2차 대사산물인 플라보노이드 합성을 조절해 항산화 효과를 높입니다.

지베렐린은 테르펜류의 생합성에 관여해 향기 물질이나 방어 물질 생산을 촉진합니다.

시토키닌은 폴리페놀 합성을 도와 노화 방지에 기여하고,
앱시스산은 스트레스 반응으로 인해 생성되는 다양한 2차 대사산물의 생산을 조절합니다.
이렇게 파이토호르몬은 식물의 2차 대사산물과 상호작용하여 식물의 성장과 생존에 중요한 역할을 합니다.
Biotechnol Rep (Amst)
. 2022 Jun 8;35:e00748. doi: 10.1016/j.btre.2022.e00748
The bioactive potential of phytohormones: A review
Arpan Mukherjee a, Anand Kumar Gaurav a, Saurabh Singh a, Shweta Yadav a, Shiuly Bhowmick a, Saman Abeysinghe b, Jay Prakash Verma a,⁎
- Author information
- Article notes
- Copyright and License information
PMCID: PMC9204661 PMID: 35719852
Highlights
- •
- •
- •
- •
- •
- 식물호르몬은 식물, 인간 및 미생물에게 생물학적 활성 물질로 작용합니다.
- •사이토키닌, GA 및 옥신은 활성산소를 감소시켜 암 및 종양 질환을 예방합니다.
- •식물호르몬은 인간용 의약품 및 화장품에 사용됩니다.
- •미생물은 식물 호르몬 생산의 잠재적 원천이 될 수 있습니다.
- •식물호르몬은 식물-동물-미생물 상호작용에서의 신호 전달에 핵심적인 역할을 합니다.
Keywords: Plant hormones, Microbes, Cell growth regulation, Disease, Human health
Abstract
Plant hormones play an important role in growth, defence and plants productivity and there are several studies on their effects on plants. However, their role in humans and animals is limitedly studied. Recent studies suggest that plant hormone also works in mammalian systems, and have the potential to reduce human diseases such as cancer, diabetes, and also improve cell growth. Plant hormones such as indole-3-acetic acid (IAA) works as an antitumor, anti-cancer agent, gibberellins help in apoptosis, abscisic acid (ABA) as antidepressant compounds and regulation of glucose homeostasis whereas cytokinin works as an anti-ageing compound. The main aim of this review is to explore and correlate the relation of plant hormones and their important roles in animals, microbes and plants, and their interrelationships, emphasizing mainly human health. The most important and well-known plant hormones e.g., IAA, gibberellins, ABA, cytokinin and ethylene have been selected in this review to explore their effects on humans and animals.
식물 호르몬은
성장, 방어 및 식물 생산성에 중요한 역할을 하며,
그 효과에 대한 연구가 다수 진행되어 왔습니다.
그러나 인간과 동물에서의 역할은 제한적으로 연구되었습니다.
최근 연구들은 식물 호르몬이 포유류 시스템에서도 작용하며,
암, 당뇨병과 같은 인간 질병을 감소시키고
세포 성장 개선에 잠재력을 가지고 있음을 제시하고 있습니다.
식물 호르몬 중 인돌-3-아세트산(IAA)은 항종양, 항암제로 작용하며,
지베렐린은 세포 사멸을 촉진하고,
아브시스산(ABA)은 항우울제 성분으로 작용하며 포도당 균형을 조절합니다.
반면 사이토키닌은 항노화 성분으로 작용합니다.
이 리뷰의 주요 목적은 식물 호르몬과 동물, 미생물, 식물 간의 관계 및 상호작용을 탐구하고,
특히 인간 건강에 초점을 맞춰 그 중요성을 규명하는 것입니다.
이 리뷰에서는
인간과 동물에 미치는 영향을 탐구하기 위해 가장 중요하고 잘 알려진 식물 호르몬인
IAA, gibberellins, ABA, cytokinin 및 ethylene이 선정되었습니다.
1. Introduction
Hormones are generally defined as the class of signalling molecules that are released or secreted from specific glands in organisms and are transported by the circulatory system to the specified targeted organs to regulate physiology and behaviour [1]. The term hormones are sometimes extended to include some important chemicals compounds produced by specified cells that affect same or other cells [2]. In the specified targeted cells, hormones bind to a specific receptor, resulting in a change in the cell function and the activation of the signal transduction pathway inside the cells [3]. In the plant cell, there are no specific hormone-secretion glands, but there are some small signals molecules/compounds produced within the plants in extremely low concentrations called hormones. These regulate the specific cellular processes in the certain targeted plant cells, with the site of function varying from the production area to other organs where they have to function [4].
Our main emphasis on five primary plant hormones that were well studied include auxins, gibberellins, cytokinin's, abscisic acid and ethylene [5]. Some chemical analogues of plant hormones work differently in different systems, such as an auxin-like chemical compound, 2,4-D or 2,4-dichlorophenoxyacetic acid, was developed to work as an herbicide in the USA (1945), which was the earliest plant growth regulatory pesticide product [6]. Other plant hormones like gibberellins, cytokinin's, and ethylene are also available in the market as a plant growth regulatory component. Plant hormones are known to work in plant growth and development. But recent study suggests that the plant hormones also work on human against different disease [7]. Synthesis of certain specific plant hormones, like, ABA, has been found not only in the plants system, but also in cyanobacteria, fungi, different animals and human beings [8]. ABA regulates animal's cell growth and differentiation, and improved the immune response in the presence of various stimuli [9]. Cytokining also help in the cell growth and development, anti-stress components [10, 11]. Auxin plays a crucial role as anti-tumour agent in human cell [12], where Gibberellin influences the antioxidant property [13]. In this review, we have attempted to explore plant hormones with the process of biosynthesis and chemical synthesis for their functional relevance in agriculture, microbial cells and the human body, and their potential applications in human health improvement.
1. 서론
호르몬은
일반적으로 유기체 내 특정 샘에서 분비되거나 방출되는 신호 분자의 한 종류로,
순환계를 통해 특정 표적 기관으로 운반되어 생리적 기능과 행동을 조절하는 물질로 정의됩니다 [1].
호르몬이라는 용어는 때로는 특정 세포에서 생성되어 동일한 세포나 다른 세포에 영향을 미치는 중요한 화학 화합물을 포함하도록 확장되기도 합니다 [2]. 특정 표적 세포에서 호르몬은 특정 수용체에 결합하여 세포 기능의 변화와 세포 내 신호 전달 경로의 활성화로 이어집니다 [3].
식물 세포에는 특정 호르몬 분비선이 없지만,
식물 내에서 극히 낮은 농도로 생성되는 작은 신호 분자/화합물이 호르몬으로 불립니다.
이들은 특정 표적 식물 세포의 특정 세포 과정을 조절하며,
기능 부위는 생산 부위에서 기능이 필요한 다른 기관으로 다양합니다 [4].
우리의 주요 연구 대상은 잘 연구된
다섯 가지 주요 식물 호르몬인
옥신, 지베렐린, 사이토키닌, 아브시스산, 에틸렌입니다 [5].
식물 호르몬의 화학 유사체는 다양한 시스템에서 다르게 작용합니다.
예를 들어, 옥신 유사 화합물인 2,4-D(2,4-디클로로페녹시초산)는 1945년 미국에서 제초제로 개발되었으며, 이는 최초의 식물 성장 조절 농약 제품입니다 [6]. 기타 식물 호르몬인 gibberellins, cytokinin, ethylene도 식물 성장 조절 성분으로 시중에 판매되고 있습니다. 식물 호르몬은 식물 성장과 발달에 작용하는 것으로 알려져 있습니다.
그러나
최근 연구는
식물 호르몬이 인간에게도 다양한 질병에 대해 작용한다는 것을 제시했습니다 [7].
특정 식물 호르몬인 ABA의 합성은
식물 시스템뿐만 아니라 시아노박테리아, 곰팡이, 다양한 동물 및 인간에서도 발견되었습니다 [8].
ABA는
동물의 세포 성장과 분화를 조절하며,
다양한 자극 하에서 면역 반응을 향상시킵니다 [9].
사이토킨은
세포 성장과 발달, 항스트레스 성분으로 작용합니다 [10, 11].
옥신은
인간 세포에서 항종양제로 중요한 역할을 합니다 [12],
반면
지베렐린은 항산화 특성에 영향을 미칩니다 [13].
이 리뷰에서는
식물 호르몬의 생합성 및 화학 합성 과정을 통해
농업, 미생물 세포 및 인간 신체에서의 기능적 관련성을 탐구했으며,
인간 건강 개선을 위한 잠재적 응용 가능성을 논의했습니다.
2. Can plant hormones affect human physiology?
Plants and microbes have shaped the whole environment of the earth for millions of years [14]. Plant hormones regulate the plant physiology, shape and associated microbial environment [15]. But some microbes show commensalism, mutualism, symbiosis, and/or harmful microbes that release and mimic some essential plant hormones to alter plant hosts and their microbial communities [15]. A very poorly appreciated fact is that animals (mainly humans) produce and are affected by plant hormones [16]. Occasionally, these plant hormones affect glucose metabolism, inflammation, antioxidant response, cellular processes, cell division, cell cycle regulation, cancer etc. (Table 1, Fig. 1) [16]. The plant hormones have a great role in human and microbial cells that are accrued either by processed diet or directly through ingestion of raw plant material. We tried to explore this poorly investigated area and illustrate it with proper examples that how plants hormones directly or indirectly affect human and microbes’ health.
2. 식물 호르몬은 인간 생리학에 영향을 미칠 수 있나요?
식물과 미생물은 수백만 년 동안 지구의 전체 환경을 형성해 왔습니다 [14]. 식물 호르몬은 식물 생리학, 형태 및 관련된 미생물 환경을 조절합니다 [15]. 그러나 일부 미생물은 공생, 상호주의, 공생 관계, 또는 유해한 미생물로 작용하며, 필수적인 식물 호르몬을 분비하거나 모방하여 식물 호스트와 그 미생물 공동체를 변화시킵니다 [15].
매우 잘 알려지지 않은 사실은
동물(주로 인간)이 식물 호르몬을 생성하고
이에 영향을 받는다는 점입니다 [16].
때로는 이러한 식물 호르몬이
포도당 대사, 염증, 항산화 반응, 세포 과정, 세포 분열, 세포 주기 조절, 암 등에
식물 호르몬은 가공된 식단을 통해 축적되거나
생식물 재료의 직접 섭취를 통해
인간과 미생물 세포에 큰 역할을 합니다.
우리는 이 잘 연구되지 않은 분야를 탐구해 보고,
식물 호르몬이 인간과 미생물의 건강에 직접적 또는 간접적으로 어떻게 영향을 미치는지
적절한 예시를 통해 설명하고자 했습니다.
Table 1.
Plant hormones and its function.
HormonesOrganismFunctionsReference
| ABA | Plants | Plant Developmental processes. Abiotic stress responses. Regulation of root growth & water conductivity. Regulation of stomatal closure under drought Condition. | [71] [69] [70] [72] |
| Humans | Glucose homoeostasis. Pro-inflammatory action in stress condition. Antidepressant property found in mice brain. Induced cell innate immunity. | [93] [78] | |
| Microbes (Endophytes) | Signalling molecules for communication amongst species. Promote interaction between plant & Arbuscular Mycorhizal fungi. | [94] [73] | |
| Cytokinin | Plants | Cell division, cell elongation and other biological Processes. Chloroplast differentiation, leaf expansion, Senescence, apical dominance, nutrient mobilization. Morphogenesis ad meristem activity. Signal transduction in stress condition. | [84] [85] |
| Humans | Gerontomodulatory and anti-ageing property. Oxidative product of DNA, affect gene expression, cell Cycle, stimulation of Ca flux anti-stress compound. Anti-oxidant property in DNA. Found in the urine of humans as a product of tryptophan metabolism in cancer patients. | [82] [83] | |
| Microbes (Endophytes) | NA | ||
| Auxin | Plants | Promote cell differentiation, vascular tissue differentiation, root tip initiation Induce ethylene production, development of seedless Fruit. Disease resistance | [47] [93] [65] |
| Humans | Auxin has anti-tumour property Regulate cell cycle | [56] | |
| Microbes | NA | ||
| Gibberellin | Plants | Stimulates shoot growth, extension of internode, flowering Stem growth, induce seed germination, fruit setting Inhibition of formation of free radical which induce lipid peroxidation. | [61] [59] |
| Humans | Causes liver and DNA damage, antioxidant defence system Antitumor property | [95] [13] | |
| Microbes | Endophytic fungus Porostereum spadiceum AGH786 Penicillium citrinum and Aspergillus fumigatus – promote plant growth | [63] [65] | |
| Ethylene | Plants | Fruit ripening, break bud and seed dormancy, regulate leaf growth and abscission of leaves | [88] |
| Humans | Ethylene oxide- genotoxic carcinogen Synthesized during oxidative stress | [90] [91] | |
| Microbes | Mucor hiemalis Defence plants from abiotic and biotic stress | [92] |
Fig. 1.
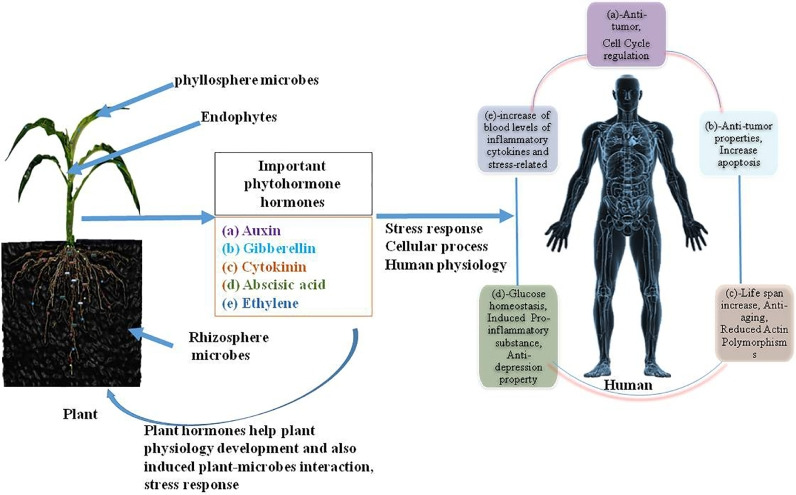
Plant hormones function for plant and human health to interlink each other.
3. Synthesis of phytohormones as bioactive compounds
3.1. Biosynthesis of phytohormones
The biosynthesis of auxin occurs via two major pathways, the first one is tryptophan dependant and the second is tryptophan independent. The flavin-containing monooxygenases tryptophan aminotransferases are known to synthesize IAA from l-tryptophan (Trp) via indole-3-pyruvate (IPA) [17]. Although, several other pathways have been identified for Trp-dependant IAA synthesis of auxin such as β-oxidation of indole-3-butyric acid (IBA) or by hydrolysing IAA conjugates, indole-3-acetamide (IAM), tryptamine (TRA), and indole-3-acetaldoxime (IAOX) pathways. Despite plants, some bacteria are also known to produce IAA by indole-3-pyruvate, indole-3-acetamide, and indole-3-acetonitrile pathways [18, 19].
The biochemical pathway for Gibberellins (GAs) synthesis involves 3 stages starting from GGPP (geranyl-geranyl diphosphate) via IPP (isopentenyl diphosphate) and is catalysed by many enzymes located in different parts of a plant cell. The first step occurs in the proplastids catalysed by enzymes which results in the formation of ent‑kaurene. Oxidation of ent‑ kaurene formed GA12-aldehyde in the second stage is then catalysed by cytochrome P-450 monooxygenases in the endoplasmic reticulum. The third stage of the pathway is catalysed by 2-oxoglutarate-dependant di-oxygenases in the cell's cytosol to form GA [20]. In fungi, GA synthesis takes place by the MVA pathway which provides IPP for the synthesis of GAs [21]. Currently, there are 136 GAs known to be produced by fungi. Some plant growth-promoting rhizobacteria are also known to produce GAs [22].
Synthesis of ABA in plants involves 2C-methyl-d-erythritol-4-phosphate (MEP) pathways that occur in plastids by the production of carotenoids and a sequential conversion of zeaxanthin into xanthoxin and finally into active ABA [23]. Whereas in fungi, ABA synthesis takes place by the mevalonic acid (MVA) pathway via farnesyl diphosphate (FPP) intermediates [24].
The biochemical reaction of ethylene synthesis occurs through the methionine dependant pathway that starts with the conversion of l-methionine into S-adenosylmethionine (SAM) and this step is ATP dependant which is catalysed by the enzyme SAM synthase. This SAM is subsequently converted into 1-aminocyclopropane −1- carboxylic acid (ACC) by the enzyme ACC synthase (ACS). The final biochemical reaction of ethylene biosynthesis is the conversion of ACC into ethylene by the presence of an enzyme ACC oxidase (ACO) [25]. Some of the pathogenic bacteria produce ethylene through the utilization of arginine and α–ketoglutarate in the presence of an ethylene-forming enzyme (EFE) [26].
The biosynthesis of cytokinin occurs via isopentenyladenine-dependant and isopentenyladenine independent pathways. In isopentenyladenine-dependant pathway, the initial process of production of tZ cytokinin is the formation of isopentenyladenine (iP) nucleotide (iPRDP/iPRTP) catalysed by adenosine phosphate isopentenyl transferase (IPT). As substrates, IPT may utilize ATP, ADP, or AMP, and as prenyl donors, dimethylallyl pyrophosphate (DMAPP) or hydroxymethylbutenyl pyrophosphate (HMBPP). Further, iPRDP/iPRTP catalysed by CYP735A into tZ nucleotide and phosphoribohydrolase convert this inactive tZ into active tZ [27,28]. In isopentenyladenine independent pathway, tZ type cytokinin formed through shifting the hydroxylated side chain of HMBDP to the adenine ring [29,30]. Cytokinin can also be synthesized in plants and bacteria through recycled tRNAs [31,32].
3. 식물 호르몬의 생합성
3.1. 식물 호르몬의 생합성
옥신(auxin)의 생합성은 두 가지 주요 경로를 통해 이루어집니다.
첫 번째 경로는 트립토판 의존적이며,
두 번째 경로는 트립토판 독립적입니다.
플라빈 함유 모노옥시게나제인 트립토판 아미노전달효소는
l-트립토판(Trp)을 통해
인돌-3-피루베이트(IPA)를 경유하여
IAA를 합성하는 것으로 알려져 있습니다[17].
그러나 트립토판 의존적 IAA 생합성을 위한 다른 경로들도 식별되었습니다.
예를 들어, 인돌-3-부티르산(IBA)의 β-산화나 IAA 공액체의 가수분해, 인돌-3-아세타미드(IAM), 트립타민(TRA), 인돌-3-아세탈도옥심(IAOX) 경로 등이 있습니다. 식물 외에도 일부 세균은 인돌-3-피루베이트, 인돌-3-아세타미드, 인돌-3-아세토니트릴 경로를 통해 IAA를 생성하는 것으로 알려져 있습니다 [18, 19].
기버렐린(GAs) 합성의 생화학적 경로는
GGPP(제라닐-제라닐 이인산)에서 시작되어
IPP(이소펜틸 이인산)를 거쳐 3단계로 진행되며,
식물 세포의 다양한 부위에 위치한 여러 효소에 의해 촉매됩니다.
첫 번째 단계는 프로플라스티드에서 효소에 의해 촉매되어 엔트-카우렌을 형성합니다. 두 번째 단계에서 엔트-카우렌의 산화는 내소체에서 사이토크롬 P-450 모노옥시게나제에 의해 촉매되어 GA12-알데히드를 생성합니다. 세 번째 단계는 세포질에서 2-옥소글루타레이트 의존성 디옥시게나제에 의해 촉매되어 GA [20]를 형성합니다. 균류에서는 GA 합성이 MVA 경로를 통해 이루어지며, 이 경로는 GA 합성에 필요한 IPP를 공급합니다 [21]. 현재 균류에 의해 생성되는 GA는 136종이 알려져 있습니다. 일부 식물 성장 촉진 뿌리 박테리아도 GA를 생성하는 것으로 알려져 있습니다 [22].
식물에서의 ABA 합성은
플라스티드에서 카로티노이드 생성과 제아잔틴에서 자탄신으로의 순차적 전환을 거쳐
최종적으로 활성 ABA로 전환되는
2C-메틸-d-에리트릴-4-인산(MEP) 경로를 통해 이루어집니다 [23].
반면
곰팡이에서는 ABA 합성이
메발론산(MVA) 경로를 통해
파르네실 디포스페이트(FPP) 중간체를 거쳐 이루어집니다 [24].
에틸렌 합성의 생화학적 반응은
메티오닌 의존적 경로를 통해 발생하며,
이는 L-메티오닌이 S-아데노실메티오닌(SAM)으로 전환되는 단계로 시작됩니다.
이 단계는 ATP에 의존하며 SAM 합성효소에 의해 촉매됩니다.
이 SAM은 이후 ACC 합성효소(ACS)에 의해 1-아미노사이클로프로판-1-카르복실산(ACC)으로 전환됩니다.
에틸렌 생합성의 최종 생화학적 반응은
ACC가 ACC 산화효소(ACO)의 존재 하에
에틸렌으로 전환되는 과정입니다 [25].
일부 병원성 세균은 에틸렌 형성 효소(EFE)의 존재 하에 아르기닌과 α-케토글루타레이트를 이용해 에틸렌을 생성합니다 [26].
사이토키닌의 생합성은
이소펜텐일아데닌 의존적 및 이소펜텐일아데닌 독립적 경로를 통해 발생합니다.
이소펜텐일아데닌 의존적 경로에서 tZ 사이토키닌의 초기 생산 과정은 아데노신 인산 이소펜텐일 전이효소(IPT)에 의해 촉매되는 이소펜텐일아데닌(iP) 핵산(iPRDP/iPRTP)의 형성입니다. IPT의 기질로는 ATP, ADP, 또는 AMP가 사용되며, 프렌일 기증체로는 디메틸알릴 피로인산(DMAPP) 또는 하이드록시메틸부텐일 피로인산(HMBPP)이 사용됩니다. 또한, CYP735A에 의해 촉매된 iPRDP/iPRTP는 tZ 핵산을 생성하며, 포스포리보히드롤라제는 이 비활성 tZ를 활성 tZ로 전환합니다 [27,28]. 이소펜텐일아데닌 독립 경로에서는 HMBDP의 수산화 측쇄를 아데닌 고리로 이동시켜 tZ 유형의 사이토키닌이 형성됩니다 [29,30]. 사이토키닌은 식물과 세균에서 재활용된 tRNA를 통해 합성될 수도 있습니다 [31,32].
3.2. Chemical synthesis of phytohormones
Some synthetic compounds share chemical and structural analogies with naturally-occurring phytohormones. Boessneck for the first time evolved the mechanism for the synthesis of 1-naphthyl-acetic acid from naphthoyl chloride and 1-methylnaphthalene. Since then many mechanisms have been developed to obtain auxin-like compounds [33]. Savaldi-Goldstein [34] identified some synthetic analogues of 2,4-dichlorophenoxyacetic acid (2,4-D), and 1-naphthaleneacetic acid (1-NAA) and termed them “Pro-auxins”. These compounds can easily diffuse to the hypocotyl of the seedlings and undergo cleavage and hydrolysis for releasing auxins.
Kinetin was first synthesized in the laboratory by Miller and his co-workers in three different ways. First, it was isolated from herring sperm DNA by autoclaving it and five times extracted with ethyl ether. Second, direct alkylation of adenine with furfuryl chloride resulted in the formation of 6-furfurylaminopurine. Another experiment was performed by heating 6-methylmercaptopurine with furfuryl amine at high temperature and then at low temperature for many hours to obtain kinetin [35]. In the same way, Okumura et al. [36] synthesized BAP (6-benzylaminopurine) by condensing 6-methylmercaptopurine with 2–3 molecular equivalents of corresponding amines. In a study, the heterocyclic chemical derivatives of cytokinin such as 6-(3-hydroxybenzylamino) purine (meta-topolin) and 6-(2-hydroxybenzylamino) purine (ortho-topolin) along with their 9-glucosides and 9-ribosides were produced through the condensation of 6- chloropurine and its 9-glycosides with the addition of hydroxybenzylamine [37]. In another study, analogues of BAP were synthesized with various substituents attached to the phenyl ring [38] and adenine moiety was substituted at N1, C2, N3, N6, N7, C8 and N9 positions [39].
A group of synthetic analogues of ABA were synthesized and found more active than naturally occurring ABA that can be used as potential plant growth regulators viz., 2′,3′-benzoabscisic acid, pyrabactin, 2,3-cyclopanated and isoabscisic acid. 2′,3′-benzoabscisic acid was synthesized by treatment of 1-tetralones with MeI in the presence of NaH to introduce germinal methyl group to the adjacent carbonyl carbon to get (Z)−3-methylpent-2-en-4-yn-1-ol and n-butyllithium and other intermediates and finally these primary alcohols are oxidized into 2′,3′-benzoabscisic acid [40, 41].
Artificially synthesized chemical compounds structurally similar to ethylene are also produced. For example, ethephon (2-Chloroethylphosphonic acid) is a synthetic ethylene used to promote flower initiation, abscission, and fruit ripening. Ethephon is chemically synthesized through the acid hydrolysis of bis(2-chloroethyl)−2-chloroethylphosphonate mediated by hydrochloric acid (HCl). However, some of the other modified pathways for ethephon synthesis have been described by the Cauret [42]. Zhang & Wen [43] also reported the synthesis of ethylene gas by the decomposition of ethephon in a disodium hydrogen phosphate-buffered solution.
Yang et al. [44] synthesized 30 chemical derivatives of thiourea having gibberellin like activities. In general, these were derived through the substituted aniline (for example, 3-(trifluoromethyl) aniline) which was used to generate aromatic isothiocyanate compounds and then reacted with a substituted aromatic ethylamine to produce a thiourea derivative having two aromatic rings. However, some other chemical compounds are also synthesized such as AC94377 (1-(3-chlorophthalimido)-cyclohexanecarboxamide) [45] and 67D (l ((S)−3-phenyl-2-(9,10-dihydro-9,10-ethanoanthracene-11,12-dicarboximido) propanoic acid) [46] identified as GA, mimic and perform same function.
3.2. 식물 호르몬의 화학 합성
일부 합성 화합물은 자연에 존재하는 식물 호르몬과 화학적 및 구조적 유사성을 공유합니다. Boessneck은 처음으로 나프토일 클로라이드와 1-메틸나프탈렌으로부터 1-나프틸아세트산(2,4-D)의 합성 메커니즘을 처음으로 규명했습니다. 이후 많은 메커니즘이 개발되어 auxin 유사 화합물을 얻는 데 활용되었습니다 [33]. Savaldi-Goldstein [34]는 2,4-dichlorophenoxyacetic acid (2,4-D)와 1-naphthaleneacetic acid (1-NAA)의 합성 유사체를 식별하고 이를 “Pro-auxins”라고 명명했습니다. 이 화합물은 묘의 hypocotyl로 쉽게 확산되어 분해 및 가수분해를 통해 auxin을 방출합니다.
키네틴은 밀러와 그의 동료들에 의해 실험실에서 세 가지 다른 방법으로 처음 합성되었습니다. 첫째, 청어 정자 DNA를 오토클레이브 처리한 후 에틸 에테르로 5회 추출하여 분리되었습니다. 둘째, 아데닌에 푸르푸릴 클로라이드로 직접 알킬화하여 6-푸르푸릴아미노푸린을 형성했습니다. 또 다른 실험에서는 6-메틸메르카토푸린을 푸르푸릴 아민과 고온에서 가열한 후 저온에서 수 시간 동안 반응시켜 키네틴을 얻었습니다 [35]. 동일한 방법으로 오쿠무라 등 [36]은 6-벤질아미노푸린(BAP)을 6-메틸메르카토푸린과 대응하는 아민 2–3 분자 당량과 축합시켜 합성했습니다. 한 연구에서 사이토키닌의 이환환식 화합물인 6-(3-하이드록시벤질아미노)푸린(메타-토폴린)과 6-(2-하이드록시벤질아미노)푸린(오르토-토폴린) 및 그 9-글루코사이드와 9-리보사이드가 6-클로로푸린과 그 9-글루코사이드에 하이드록시벤질아민을 추가하여 축합시켜 생성되었습니다 [37]. 다른 연구에서는 페닐 고리에 다양한 치환기가 부착된 BAP의 아날로그가 합성되었으며, 아데닌 모티프는 N1, C2, N3, N6, N7, C8 및 N9 위치에 치환되었습니다. [39].
ABA의 합성 유사체 그룹이 합성되었으며, 자연적으로 발생하는 ABA보다 더 활성도가 높아 식물 성장 조절제로 사용될 수 있는 2′,3′-벤조아브시시산, 피라바크틴, 2,3-사이클로판화 및 이소아브시시산이 발견되었습니다. 2′,3′-벤조아브시시산은 1-테트라론에 NaH 존재 하에 MeI로 처리하여 인접한 카르보닐 탄소에 발아 메틸 그룹을 도입하여 (Z)-3-메틸펜트-2-엔-4-인-1-올과 n-부틸리튬 및 기타 중간체를 생성한 후, 최종적으로 이 일차 알코올을 산화시켜 2′,3′-벤조아브시시산을 합성했습니다 [40 , 41].
에틸렌과 구조적으로 유사한 인공 합성 화합물도 생산됩니다. 예를 들어, 에테폰(2-클로로에틸포스폰산)은 꽃 개화, 탈락, 과일 숙성을 촉진하기 위해 사용되는 합성 에틸렌입니다. 에테폰은 염산(HCl)을 매개로 한 bis(2-클로로에틸)−2-클로로에틸포스폰산의 산 가수분해로 화학적으로 합성됩니다. 그러나 에테폰 합성의 다른 변형 경로는 Cauret [42]에 의해 설명되었습니다. Zhang & Wen [43]은 또한 디소듐 수소인산염 완충 용액에서 에테폰의 분해를 통해 에틸렌 가스를 합성했다고 보고했습니다.
Yang et al. [44]은 지베렐린 유사 활성을 가진 티우레아의 30가지 화합물 유도체를 합성했습니다. 일반적으로 이러한 화합물은 대체된 아닐린(예: 3-(트리플루오로메틸) 아닐린)을 사용하여 아로마틱 이소티오시아네이트 화합물을 생성한 후, 대체된 아로마틱 에틸아민과 반응시켜 두 개의 아로마틱 고리를 가진 티우레아 유도체를 생성하는 과정을 통해 유도되었습니다. 그러나 AC94377 (1-(3-클로로프탈이미도)-사이클로헥산카르복사미드) [45] 및 67D (l ((S)−3-페닐-2-(9,10-디히드로-9,10-에타노안트라센-11,12-디카르복시미도) 프로파노산) [46]는 GA와 유사하며 동일한 기능을 수행하는 것으로 확인되었습니다.
4. Plant hormones and their effects on human health
4.1. Auxin
Auxin is an important phytohormone, that performs a lot of functions in the plant including plant cell division, enlargement of a cell, stimulation of the vascular tissue differentiation, root initiation, tropic response and also delay in leaf senescence (Table 1, Fig. 1) [1]. Normally, IAA is produced from the apical meristematic region of plants shoots, buds and the tip of the root. The effect of auxins in plants is dependant on their concentration. The higher amount of auxin inhibits root elongation and vice versa [47]. It has been also reported that auxin synergistically interacts with ethylene, and leads to the development of the processes of root hair formation and root elongation, where the antagonistic activity shows hypocotyl elongation and lateral root formation in plants [48]. Gustafson in 1937 [49], for the first time, demonstrated that the application of auxins like components into the stigma of tomato and many other plants species causes the development of parthenocarpic fruit. During anthesis or probably after pollination and fertilization, the level of auxin and gibberellins consistently increased resulting in the development of seedless fruit. Auxin can induce biosynthesis of gibberellins and it can increase auxin levels in the ovary of plants [50]. It was reported that during plant defence, the auxin pathway behaves antagonistically with SA (Salicylic acid) pathway whereas the auxin and JA (Jasmonic acid) signalling pathway interact positively with each other [51]. Auxin and JA signalling pathways, share commonalities in providing resistance to necrotrophic pathogens. TIR1 and COI1 (F-box protein for Auxin and JA) get involved in providing resistance to several necrotrophic pathogens [52].
Auxin is also found in some parts of the mammalian body, mainly synthesized from an essential amino acid tryptophan which is consumed from different kinds of vegetables (soybean, broccoli, beans, onion and leafy vegetables) and other animals products, such as milk, cheese, red meat, chickpea, pumpkin seed, almonds and peanut (https://www.myfooddata.com/articles/high-tryptophan-foods.php). From a few previous studies, it was known that mammalian cells’ function can be influenced by auxins. It was reported that auxin has a potential role to work as an antitumor agent [53]. Some recent scientific studies reported that in mammals, delay in cell cycle (S, G2 or M) progression in cells happened due to suppression of SCF typeubiquitin ligase function (by suppressing SKP2 dependant CDK2 and CDK4 activity) which induces G1 cell cycle arrest. For example, in ovarian cancer cells of humans, G1 arrest is caused by suppressing SKP2 which is the f-box protein of SCF complex and control proteolysis of CDK inhibitors including p27 and p21. Suppression of SKP2 reduced the progress of the G1 cell cycle and decreases the percentage of cells in S-phase with the help of auxin [53] (Table 1, Figs. 1 and 2).
4. 식물 호르몬과 인간 건강에 미치는 영향
4.1. 옥신
옥신은
식물 세포 분열, 세포 크기 증가, 혈관 조직 분화 촉진, 뿌리 형성, 방향 반응 및 잎 노화 지연 등
식물에서 다양한 기능을 수행하는 중요한 식물 호르몬입니다(표 1, 그림 1) [1].
일반적으로 IAA는 식물 줄기, 싹 및 뿌리 끝의 정점 분열 조직에서 생성됩니다. 옥신의 식물 내 효과는 농도에 따라 달라집니다. 옥신의 농도가 높을수록 뿌리 연장 억제 효과가 나타나며, 반대로 농도가 낮을수록 연장 효과가 증가합니다 [47]. 또한 옥신은 에틸렌과 시너지 효과를 발휘하여 뿌리 털 형성 및 뿌리 연장 과정의 발달을 촉진하며, 이 과정에서 대항 작용은 hypocotyl 연장 및 측근 형성을 유발합니다 [48].
Gustafson은 1937년 [49]에서 처음으로 토마토와 다른 많은 식물 종의 암술에 auxin 유사 성분을 적용하면 무수정 결실(parthenocarpic fruit)이 발생함을 처음으로 입증했습니다. 꽃피기 기간이나 아마도 수분 및 수정 후, auxin과 gibberellin의 수준이 지속적으로 증가하여 씨 없는 과일이 형성되었습니다. 옥신은 gibberellin의 생합성을 유도하며, 식물의 난소 내 옥신 농도를 증가시킵니다 [50]. 식물 방어 과정에서 옥신 경로는 SA (살리실산) 경로와 대항적으로 작용하지만, 옥신과 JA (자스모닉산) 신호 경로는 서로 긍정적으로 상호작용합니다 [51]. 옥신과 JA 신호 경로는 necrotrophic 병원체에 대한 저항성을 제공하는 데 공통점을 공유합니다. TIR1과 COI1(옥신과 JA를 인식하는 F-박스 단백질)은 여러 괴사성 병원체에 대한 저항성을 제공하는 데 관여합니다 [52].
옥신은
포유류의 일부 조직에서도 발견되며,
주로 필수 아미노산인 트립토판에서 합성됩니다.
트립토판은
다양한 채소(콩, 브로콜리, 콩류, 양파, 잎채소)와
동물성 제품(우유, 치즈, 붉은 고기, 병아리콩, 호박씨, 아몬드, 땅콩)에서
섭취됩니다 (https://www.myfooddata.com/articles/high-tryptophan-foods.php).
이전 연구에서 포유류 세포의 기능이 옥신에 의해 영향을 받을 수 있다는 것이 알려져 있습니다. 옥신이 항종양제로 작용할 잠재적 역할을 가질 수 있다는 보고가 있습니다 [53].
최근 과학적 연구에서는
포유류에서 세포 주기(S, G2 또는 M) 진행 지연이
SCF 유형 유비퀴틴 리가제 기능 억제(SKP2에 의존하는 CDK2 및 CDK4 활성 억제)로 인해 발생하며,
이는 G1 세포 주기 정지를 유발한다는 것이 보고되었습니다.
예를 들어, 인간 난소 암 세포에서 G1 정지는 SCF 복합체의 f-box 단백질인 SKP2를 억제함으로써 발생하며, 이는 p27 및 p21을 포함한 CDK 억제제의 분해를 조절합니다. SKP2의 억제는 auxin의 도움으로 G1 세포 주기 진행을 감소시키고 S-상 세포의 비율을 감소시킵니다 [53] (표 1, 그림 1 및 2).
Fig. 2.
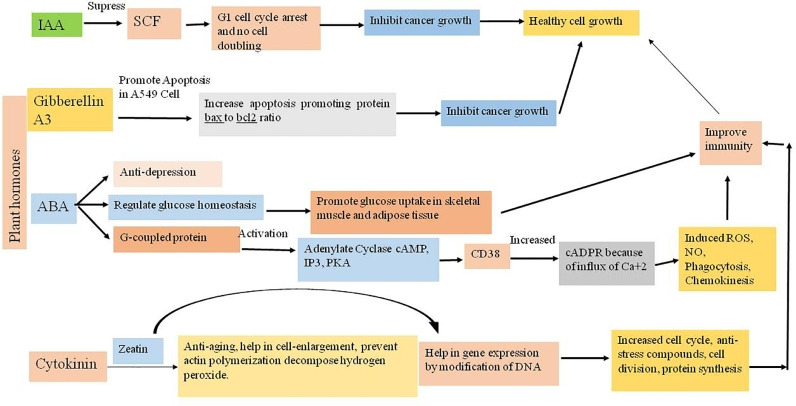
Plant hormones and its application for modulation the biochemical and functional properties of human body.
Some microorganisms are reported that were able to produce auxin hormones. IAA producing different bacterial genera include Pseudomonas, Azospirillum, Enterobacter, Streptomyces, Pantoea, Rhizobium,
Azotobacter and Alcaligenes [54], [55], [56] (Fig. 3). In various plants, microorganisms present in the rhizosphere can synthesize IAA as the secondary metabolite. For commercial IAA production, Streptomyces griseoviridis K61 and Streptomyces lydicus WYEC108 were used under trade name Mycostop [57]. Mainly IAA is produced by the microorganism helps plants directly or indirectly, consequently affecting humans (Fig. 1).
식물 호르몬 및 인간 신체 생화학적 및 기능적 특성 조절을 위한 적용.
일부 미생물이 옥신 호르몬을 생산할 수 있다는 보고가 있습니다. IAA를 생산하는 다양한 세균 속에는 Pseudomonas, Azospirillum, Enterobacter, Streptomyces, Pantoea, Rhizobium,
Azotobacter 및 Alcaligenes [54], [55], [56] (그림 3)가 포함됩니다. 다양한 식물에서 뿌리권에 존재하는 미생물은 IAA를 2차 대사산물로 합성할 수 있습니다. 상업용 IAA 생산을 위해 Streptomyces griseoviridis K61과 Streptomyces lydicus WYEC108이 Mycostop [57]이라는 상표명으로 사용되었습니다. 주로 미생물이 생산하는 IAA는 식물에 직접적 또는 간접적으로 도움을 주며, 결국 인간에게 영향을 미칩니다 (그림 1).
Fig. 3.
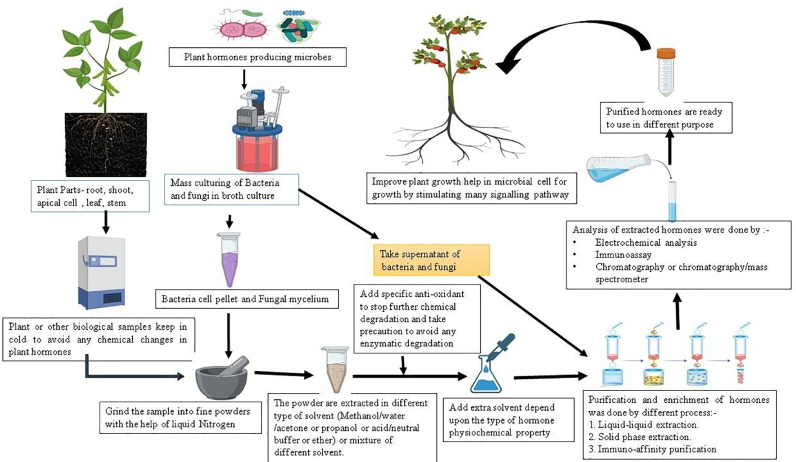
Diagrammatic representation of plant hormones extraction and purification from plant and microbes.
4.2. Gibberellins (GAs)
Gibberellins are also one of the most important phytohormones involved in plant growth promotion by degrading the negative growth regulator protein called DELLA [58] GA is known as a strong accelerator for shoot growth but shows very little response on root growth. To combat against different environmental stress conditions, various types of phytohormones are produced by plants that increase the potential to grow in such conditions. Gibberellin is one of the phytohormones secreted during stress conditions. The increased levels of GA3 were found in wheat and rice and enhanced growth under saline conditions [59]. GAs involved mainly in stem growth, induce seed germination by producing enzymes during seed germination and help in fruit setting and growth [1]. It was noticed that GAs inhibited the formation of free radicals which induce lipid peroxidation [58]. The most noticeable effect of GAs in plant growth was the extension of internode in shoot growth and enhanced apical dominance and increased leaf growth. An increased amount of GAs has very little effect on a plant which shows increased dry weight. This indicated the secondary effect of GA on leaf growth. This secondary effect was mainly due to increase in carbon fixation [60]. Very limited studies have been done on the regulation of GAs in root growth. Externally applied GAs showed very little effect on the growth of roots in various plants. In several controlling developmental processes such as germination, growth and flowering, GAs interacts antagonistically with ABA [61].
Gibberellic acid in the human body comes from a plant diet (vegetables and fruits treated with gibberellic acid and some vegetables naturally produce gibberellic acids such as chilli and olive). The metabolic processes of humans can be influenced by the phytohormones present in our diet but the effect of GAs on humans is very less reported. GAs was reported as a potent pro-oxidant, that induce oxidative stress by generating reactive oxygen species (suppress the function of antioxidant enzymes activities) and causes hepatotoxicity in adult male albino rats by lipid peroxidation in the liver cells [62]. Cytogenetic effect of gibberellin A3 at different concentrations was noted in human lymphocyte culture. Gibberellin A3 induced chromosomal changes (by gap deletion, break), exchange between sister chromatids and DNA damage [63]. Zhang et al. [64] showed that a gibberellin derivative (GA13315) has an antitumor property against tumour cell lines in vivo as well as in vitro. GA13315 promoted apoptosis in A549 cells by increasing the level of apoptosis promoting protein (bax to bcl2 protein) ratio (Table 1, Figs. 1 and 2).
A mutualistic relationship is formed by rhizospheric bacteria with host plant, provides several benefits to them by siderophore production, increase availability of nutrient, nitrogen fixation, phosphate solubilisation and secretion of phytohormone. The different bacterial genera like Pseudomonas, Bacillus, Azotobacter, Flavobacterium, Micrococcus, Rhizobium, Xanthomonas, Agrobacterium, Clostridium and Azospirillum enhances plant growth and supresses phytopathogens [65] (Fig. 3). Endophytic microorganisms can produce phytohormones which help plants to alleviate various limiting factors such as salt stress and nutrition deficiency. It was found by Hamayun et al. [66] that an endophytic fungus Porostereum spadiceum AGH786 has the potential to produce GA which helps to alleviate salt stress and promote growth and health benefits in soyabean plant. Some of the endophytic fungi Penicillium citrinum and Aspergillus fumigatus have been reported to produce gibberellin in the rhizosphere and promote plant growth [67] (Fig. 3). Joo et al. [65] isolated a bacterial strain identified as Burkholderia sp. KCTC 11096BP through phylogenetic analysis 16S rDNA, produces GA and promote growth in cucumber and crown daisy. It has been reported that most pathogenic fungi and bacteria used GA as a secondary metabolite and this helps in plant-microbe interaction.
4.2. 지베렐린 (GAs)
지베렐린은 식물 성장 촉진에 관여하는 가장 중요한 식물 호르몬 중 하나로, 부정적 성장 조절 단백질인 DELLA [58]를 분해함으로써 작용합니다. GA는 줄기 성장의 강력한 촉진제로 알려져 있지만 뿌리 성장에는 거의 반응하지 않습니다. 다양한 환경 스트레스 조건에 대응하기 위해 식물은 이러한 조건에서 성장 잠재력을 높이는 다양한 유형의 식물 호르몬을 생성합니다. 기베렐린은 스트레스 조건에서 분비되는 식물 호르몬 중 하나입니다. GA3의 농도가 밀과 쌀에서 증가했으며 염분 조건 하에서 성장이 촉진되었습니다 [59]. GAs는 주로 줄기 성장에 관여하며, 종자 발아 시 효소를 생성하여 종자 발아를 유도하고 과실 형성 및 성장에 도움을 줍니다 [1]. GAs는 지질 과산화를 유발하는 자유 라디칼의 형성을 억제한다는 것이 관찰되었습니다 [58]. 식물 성장에서 GAs의 가장 눈에 띄는 효과는 줄기 성장 시 절간 연장, 정점 우세 강화, 잎 성장 증가입니다. GAs의 증가량은 건조 중량이 증가한 식물에 거의 영향을 미치지 않았습니다. 이는 GA의 잎 성장에 대한 2차 효과를 나타냅니다. 이 2차 효과는 주로 탄소 고정 증가 [60]에 기인합니다. 뿌리 성장에서의 GAs 조절에 대한 연구는 매우 제한적입니다. 외부적으로 적용된 GAs는 다양한 식물의 뿌리 성장에 거의 영향을 미치지 않았습니다. 발아, 성장, 꽃 피기 등 여러 발달 과정의 조절 과정에서 GAs는 ABA와 [61]와 같은 방식으로 상호작용합니다.
인체 내의 gibberellic acid는 식물성 식단(gibberellic acid로 처리된 채소와 과일, 고추와 올리브와 같은 일부 채소는 자연적으로 gibberellic acid를 생성합니다)에서 유래합니다. 인체의 대사 과정은 식이 중의 식물 호르몬에 의해 영향을 받을 수 있지만, GA가 인간에 미치는 영향은 매우 적게 보고되었습니다. GA는 강력한 프로옥시던트로 보고되었으며, 활성산소종(ROS)을 생성하여 항산화 효소 활성을 억제함으로써 산화 스트레스를 유발하며, 성인 수컷 알비노 쥐의 간 세포에서 지질 과산화 작용을 통해 간독성을 유발합니다 [62]. 기버렐린 A3의 다양한 농도에서 인간 림프구 배양에서 세포유전학적 효과가 관찰되었습니다. 기버렐린 A3는 염색체 변화(간격 삭제, 파열), 자매 염색체 간 교환 및 DNA 손상을 유발했습니다 [63]. Zhang 등 [64]는 기버렐린 유도체(GA13315)가 체내 및 체외에서 종양 세포 라인에 대한 항종양 효과를 나타냈다고 보고했습니다. GA13315는 A549 세포에서 세포 사멸 촉진 단백질(bax 대 bcl2 단백질) 비율을 증가시켜 세포 사멸을 촉진했습니다(표 1, 그림 1 및 2).
뿌리권 세균은 숙주 식물과 공생 관계를 형성하여 siderophore 생산, 영양소 가용성 증가, 질소 고정, 인산염 용해 및 식물 호르몬 분비 등을 통해 숙주 식물에 다양한 혜택을 제공합니다. Pseudomonas, Bacillus, Azotobacter, Flavobacterium, Micrococcus, Rhizobium, Xanthomonas, Agrobacterium, Clostridium 및 Azospirillum과 같은 다양한 세균 속은 식물 성장 촉진과 식물 병원균 억제 효과를 나타냅니다 [65] (그림 3). 내생 미생물은 식물이 염분 스트레스 및 영양 결핍과 같은 다양한 제한 요인을 완화하는 데 도움을 주는 식물 호르몬을 생산합니다. Hamayun 등 [66]은 내생 곰팡이 Porostereum spadiceum AGH786이 소이콩 식물의 염분 스트레스를 완화하고 성장 및 건강을 촉진하는 GA를 생산할 잠재력을 가지고 있음을 발견했습니다. 내생 곰팡이 Penicillium citrinum과 Aspergillus fumigatus는 뿌리권에서 gibberellin을 생성하여 식물 성장을 촉진한다는 보고가 있습니다 [67] (그림 3). Joo 등 [65]은 16S rDNA 계통 분석을 통해 Burkholderia sp. KCTC 11096BP로 확인된 세균 균주를 분리했으며, 이 균주는 GA를 생산하고 오이 및 크라운 데이지의 성장을 촉진합니다. 대부분의 병원성 곰팡이와 세균은 GA를 2차 대사산물로 사용하며, 이는 식물-미생물 상호작용에 도움을 준다는 보고가 있습니다.
4.3. Abscisic acid (ABA)
Abscisic acid is an important phytohormone that takes part in various developmental processes such as seed and bud dormancy, regulation of opening and closing of stomata, controls size of various parts in plants, and works in environmental stress response. ABA shows an antagonistic relationship with GA in developmental processes such as seed dormancy and germination by promoting the biosynthesis of storage compounds and preventing precautious seed germination [68]. Under abiotic stress conditions, the expression of stress tolerance genes is regulated by phytohormone ABA, which enhances plant growth in an unfavourable environment [69]. It has been also reported that ABA induces root growth and control water conductivity under drought conditions [70]. ABA has a high heat tolerance capacity so that it can maintain its vitality under high temperatures and promotes seed development, vegetative growth and seed sprouting under ecological stress conditions [71]. In addition, ABA plays an important role in stomatal closure in plants under drought conditions when there was no need for CO2 intake from the environment so that plants can reduce transpiration to prevent excess water loss from the plant body [72]. Biosynthesis of ABA in plants occurred in plastids through the MEP pathway by using carotenoid as a precursor molecule [73, 68]. It involves a series of enzymes mediating epoxidation of zeaxanthin, isomerization and cleavage of carotenoid into ABA molecules [74]. ABA also modulates plant defence responses through transcriptional reprogramming of plant cell metabolism and by inducing the expression of catalase which scavenge H2O2 [58]. Human beings and other animals are constantly exposed to ABA by various nutritional sources. Daily intake of fruits (i.e., apple, banana, apricot, avocado, fig, citrus) and vegetables (i.e., potato, tomato, soybean, barley, maize, pea, cucumber etc.) in our diet leads to the accumulation of ABA in our body [75]. High concentration of ABA in human body can be linked with a number of physiological and metabolic responses. Many studies have reported that glucose homoeostasis in a human being is regulated by ABA [76]. The adipose tissue of human releases ABA in low as well as in high glucose concentrations. In hyperglycaemia condition, release of GLP-1 induces β cells and insulinoma cells to release ABA and insulin which promotes glucose uptake in skeletal muscles and adipocytes. GLP1R is a glucagon receptor that stimulates GLP-1 to induce pancreatic β- cells for insulin secretion. It was also reported in rat myoblasts and murine adipocytes that ABA stimulates glucose uptake [77]. ABA is also known as an endogenous pro-inflammatory substance, released by animal cells during abiotic stress. Influx of Ca2+inside the granulocyte is induced by ABA. The interaction between ABA and G-coupled proteins of plasma membrane trigger the activation of adenylate cyclase [AC], overproduction of cAMP, increase of IP3, PKA mediated stimulation of CD38, and increased production of cADPR. This mechanism may be involved in influx of Ca2+ [71]. By this series of signalling pathway ABA also behaves as a pro-inflammatory endogenous cytokine as it stimulates phagocytosis, ROS and NO production, chemotaxis and chemokinesis and thus induces cell's innate immunity [78]. ABA also present in the hypothalamus and other parts of brain of mammals such as rodents and pigs. The amount of ABA in hypothalamus was found higher than any other tissues of mammals [79]. Endogenous secretion of ABA in the brain can be correlated with the stress response activity of hypothalamus. Antidepressant property of ABA was also reported in mice [79]. Thus, ABA rich food in our daily life could help to increase our innate immune responses and can also prevent different metabolic disorders like type 2 diabetes (Figs. 1 and 2).
4.3. 아브시스산 (ABA)
아브시스산은 종자 및 싹의 휴면, 기공의 개폐 조절, 식물 각 부위의 크기 조절, 환경 스트레스 반응 등에 참여하는 중요한 식물 호르몬입니다. ABA는 종자 휴면 및 발아와 같은 발달 과정에서 GA와 대립 관계를 보여 저장 물질의 생합성을 촉진하고 예방적 종자 발아를 방지합니다 [68]. 비생물적 스트레스 조건 하에서 식물 호르몬 ABA는 스트레스 내성 유전자 발현을 조절하여 불리한 환경에서 식물 성장을 촉진합니다 [69]. 또한 ABA는 가뭄 조건 하에서 뿌리 성장과 수분 전도도를 조절한다는 보고도 있습니다 [70]. ABA는 높은 열 내성 능력을 갖추어 고온 조건에서도 생명을 유지하며, 생태적 스트레스 조건 하에서 종자 발달, 생장 및 발아를 촉진합니다 [71]. 또한 ABA는 환경으로부터 이산화탄소(CO₂) 흡수가 필요 없는 가뭄 조건에서 식물의 기공 폐쇄에 중요한 역할을 하여 식물이 과도한 수분 손실을 방지하기 위해 증발을 줄일 수 있도록 합니다 [72]. 식물에서의 ABA 생합성은 카로티노이드를 전구체 분자로 사용하여 MEP 경로를 통해 플라스티드에서 발생합니다 [73, 68]. 이 과정은 제아잔틴의 에폭시화, 카로티노이드의 이성질화 및 분해 과정을 통해 ABA 분자로 전환되는 일련의 효소 반응을 포함합니다 [74]. ABA는 식물 세포 대사 과정의 전사 재프로그래밍을 통해 식물 방어 반응을 조절하며, H₂O₂를 제거하는 카탈라제의 발현을 유도합니다 [58].
인간을 포함한 동물은
다양한 영양소 원천을 통해 지속적으로 ABA에 노출됩니다.
우리 식단에
과일(예: 사과, 바나나, 복숭아, 아보카도, 무화과, 시트러스)과
채소(예: 감자, 토마토, 콩, 보리, 옥수수, 완두콩, 오이 등)를 매일 섭취하면
체내에 ABA가 축적됩니다 [75].
인간 체내의 ABA 고농도는 다양한 생리적 및 대사 반응과 연관될 수 있습니다. 많은 연구에서 인간에서의 포도당 균형이 ABA에 의해 조절된다는 것이 보고되었습니다 [76]. 인간 지방 조직은 저혈당 및 고혈당 조건에서 ABA를 방출합니다. 고혈당 상태에서 GLP-1의 방출은 β 세포와 인슐린 세포가 ABA와 인슐린을 방출하도록 유도하여 골격근과 지방 세포에서의 포도당 흡수를 촉진합니다. GLP1R은 GLP-1을 자극하여 췌장 β 세포의 인슐린 분비를 유도하는 글루카곤 수용체입니다. 쥐의 근육 전구세포와 쥐의 지방세포에서 ABA가 포도당 흡수를 촉진한다는 보고도 있습니다 [77]. ABA는 비생물적 스트레스 시 동물 세포에서 분비되는 내인성 프로염증 물질로 알려져 있습니다. ABA는 과립구 내 Ca2+ 유입을 유도합니다. ABA와 세포막의 G-결합 단백질 간의 상호작용은 아데닐산 사이클레이즈(AC) 활성화, cAMP 과다 생산, IP3 증가, PKA 매개 CD38 자극, cADPR 생산 증가를 유발합니다. 이 메커니즘은 Ca2+ 유입과 관련될 수 있습니다 [71]. 이 신호 전달 경로를 통해 ABA는 염증 촉진 내인성 사이토킨으로 작용하여 식작용, ROS 및 NO 생성, 화학유동 및 화학유동성 자극을 통해 세포의 선천적 면역 반응을 유도합니다 [78]. ABA는 설치류와 돼지 등 포유류의 뇌 시상하부 및 기타 부위에서도 발견됩니다. 뇌하수체 내 ABA의 양은 포유류의 다른 조직보다 높게 측정되었습니다 [79]. 뇌 내 ABA의 내인성 분비는 뇌하수체의 스트레스 반응 활동과 관련될 수 있습니다. ABA의 항우울제 효과는 쥐에서 보고되었습니다 [79]. 따라서 일상 생활에서 ABA가 풍부한 식품은 우리 몸의 선천적 면역 반응을 강화하고 제2형 당뇨병과 같은 다양한 대사 장애를 예방하는 데 도움이 될 수 있습니다 (그림 1 및 2).
4.4. Cytokinin
Cytokinin is a known phytohormone that occurs naturally as well as synthetically in the environment. Cytokinin is found naturally in plants and plays a vital role in cell division, differentiation and a number of biological processes such as apical dominance, leaf expansion and senescence, nutrient mobilization, chloroplast differentiation and activation of shoot meristem [80]. Deficiency of cytokine causes stunted shoot and smaller apical meristem. It is also found that cytokinin is important for morphogenesis and plant meristem activity in plants. In plants cytokinin plays an important role in signal transduction for different environmental stresses such as drought, salinity, and temperature via signalling pathways [81]. Cytokinin signalling pathway is regulated by His-Asp-phosphor lay; these components transmit the signal which triggers the CK responsive gene [81].
Other than plants, cytokinin is also found in different animals. At a very first, it was reported in an autoclaved herring sperm by Miller [82]. In mammals, cytokinin affects various physiological activities. Voller [83] discovered that cytokinin and their derivatives increased the lifespan in Caenorhabditis elegans. Zeatin, [6-[4‑hydroxy-3-methyl‑but-2-enylamino] adenine], a known cytokinin plant growth factor showed geronto-modulatory and anti-ageing properties in human skin fibroblasts in vitro. It has been experimented that long term treatment of zeatin in human fibroblasts can preserve cell vitality by reducing intracellular debris, cell enlargement, preventing actins polymerization and also by increasing the cell's ability to decompose hydrogen peroxide [84]. It has been reported that kinins are found in DNA as a product of oxidative, secondary modification and a secondary reaction of DNA, affecting different biological activities such as gene expression, inhibition of auxin action, stimulation of calcium flux, cell-cycle and as an anti-stress compound. It also induced cell division, cell differentiation and even protein synthesis in mammals [85]. Barciszewski, et al. [85] reported that high concentrations of kinins have been found in the urine of patients suffering from lung cancer. Cytokinins can be used as a marker for DNA damage as it protects DNA from oxidative stress, caused due to reactive oxygen species [ROS]. Kinetin is proven to have protective activities against oxidative stress in mammalian cells such as HL60 cells, HaCaT human keratinocyte cells, NRK rat epithelial kidney cells and human peripheral lymphocytes [86]. It has been found that cytokinin can protect cells from apoptosis and a low concentration of it can reduce stress mediated cell death [86]. Barciszewski et al. [87] hypothesized that endogenous Kinetin may arise as a consequence of oxidative damage of DNA, thus creating protection near the site of damage. Cytokinin bases have been shown to promote differentiation of keratinocytes. Kinetin at 40–200 lM concentration induces growth arrest and changes of several markers of differentiation (keratin K10 and involucrin) in human keratinocytes in cell culture [88] and also reported that treatment with Kinetin improved the sensitivity of ageing keratinocytes to the differentiating effects of Ca2+ ions (Figs. 1 and 2).
4.4. 사이토키닌
사이토키닌은 자연적으로 및 인공적으로 환경에서 발견되는 알려진 식물 호르몬입니다. 사이토키닌은 식물에서 자연적으로 발견되며 세포 분열, 분화 및 정점 우세, 잎 확장, 노화, 영양분 이동, 엽록체 분화, 줄기 분열 조직의 활성화 등 다양한 생물학적 과정에 중요한 역할을 합니다. [80] 사이토키닌 결핍은 줄기 성장 저하와 정점 분열 조직의 축소를 유발합니다. 또한 사이토키닌은 식물의 형태 발생과 분열 조직 활동에 중요하다는 것이 밝혀졌습니다. 식물에서 사이토키닌은 가뭄, 염분, 온도 등 다양한 환경 스트레스에 대한 신호 전달 경로를 통해 신호 전달에 중요한 역할을 합니다[81]. 사이토키닌 신호 전달 경로는 히스티딘-아스파르트산-인산염(His-Asp-phosphor lay)에 의해 조절되며, 이 성분들은 신호를 전달하여 CK 반응 유전자를 활성화합니다[81].
식물 외에도 사이토키닌은 다양한 동물에서도 발견됩니다. 처음에는 밀러 [82]에 의해 고압 증기 처리된 청어 정자에서 보고되었습니다. 포유류에서 사이토키닌은 다양한 생리적 활동에 영향을 미칩니다. 볼러 [83]는 사이토키닌과 그 유도체가 Caenorhabditis elegans의 수명을 연장시킨다는 것을 발견했습니다. 사이토키닌 식물 성장 인자인 제아틴 [6-[4-하이드록시-3-메틸-부-2-엔일아미노] 아데닌]은 체외에서 인간 피부 섬유아세포에서 노화 조절 및 항노화 특성을 나타냈습니다. 인간 섬유아세포에서 제아틴의 장기 투여가 세포 내 잔여물 감소, 세포 비대 방지, 액틴 중합 억제, 수소 과산화물 분해 능력 증가를 통해 세포 활력을 유지한다는 실험 결과가 보고되었습니다 [84]. 키닌은 DNA의 산화적 2차 변형 및 DNA의 2차 반응의 산물로 DNA에 존재하며, 유전자 발현, 옥신 작용 억제, 칼슘 유동 촉진, 세포 주기 조절 및 항스트레스 화합물로 작용하는 다양한 생물학적 활동을 영향을 미칩니다. 또한 포유류에서 세포 분열, 세포 분화 및 단백질 합성을 유도했습니다 [85]. Barciszewski 등. [85]는 폐암 환자의 소변에서 키닌의 고농도가 발견되었다고 보고했습니다. 사이토키닌은 활성산소종(ROS)에 의해 유발되는 산화 스트레스로부터 DNA를 보호하기 때문에 DNA 손상의 지표로 사용될 수 있습니다. 키네틴은 HL60 세포, HaCaT 인간 각질세포, NRK 쥐 신장 상피세포 및 인간 말초 림프구와 같은 포유류 세포에서 산화 스트레스에 대한 보호 활동을 나타냅니다 [86]. 사이토키닌이 세포 사멸로부터 세포를 보호하며, 낮은 농도에서 스트레스 매개 세포 사멸을 감소시킨다는 것이 밝혀졌습니다 [86]. Barciszewski 등 [87]은 내인성 키네틴이 DNA의 산화 손상으로 인해 발생하여 손상 부근에서 보호 작용을 일으킬 수 있다고 가설을 제기했습니다. 사이토키닌 기반 화합물은 케라티노사이트의 분화를 촉진하는 것으로 나타났습니다. 40–200 lM 농도의 키네틴은 인간 케라티노사이트 세포 배양에서 성장 정지와 분화 표지자(케라틴 K10 및 인볼루크린)의 변화를 유도하며 [88], 또한 키네틴 처리로 노화된 케라티노사이트의 Ca2+ 이온에 의한 분화 효과에 대한 감수성이 향상되었다고 보고되었습니다 (그림 1 및 2).
4.5. Ethylene
Ethylene is the only gaseous plant hormone that helps in, seed germination, flowering, sex determination, fruit ripening and abscission in plants. The regulation of growth and senescence of leaves is dependant on the concentration of ethylene. Ethylene interacts with other phytohormones either positively or negatively. In Arabidopsis, ethylene behaves antagonistically with abscisic acid on seed dormancy. Above a certain concentration, both inhibit elongation of the root. When the ethylene is treated with an ethylene synthesis inhibitor, its content gets lowered and ABA becomes effective in inhibition of root growth, this indicates that ethylene negatively interacts with ABA in root growth response [89]. Stepanova et al. [90] found that ethylene stimulates auxin biosynthesis for root growth promotion. They showed that tryptophan production is enhanced by stimulating WEI7 (Weak Ethylene Insensitive7) and WEI2 transcription which finally leads to auxin production in seedlings of Arabidopsis. Ethylene controls the level of auxin response factor [ARF] by regulating HOOKLESS1 [a member of N-Acetyltransferase gene family] in the hypocotyl region of plants which results in a well maintained and defined differential growth pattern found in these tissues [90]. It was found that ethylene concentration increased after interacting with herbivores. It could be due to oral secretory compounds which are secreted by herbivores and microorganisms that are transferred when insects feed on plant.
Ethylene comes from the natural metabolic process and through the air in the body of a human. In gut microflora of human, ethylene is present and can produce ethylene oxide, a known genotoxic carcinogen (IARC, Group 1) for human. In exhaled air of human, ethylene is also found [91]. Laurent et al. [92] reported that during systemic inflammation in humans, ethylene is produced. In systemic inflammation and infection, reactive oxygen species [ROS] induce oxidative stress which causes endogenous lipid peroxidation of unsaturated fatty acid and leads to ethylene formation (Figs. 1 and 2). Respiratory burst is responsible for oxidative stress and it occurs when bacterial and fungal infections encounter pathogen destroying blood cells (monocytes and neutrophils). ROS is produced in high amount due to this burst and causes oxidative stress. They showed that fatty acid (docosahexaenoic acid and oleic acid) significantly produces ethylene after exposing with Fe3+ ions which causes oxidative stress and leads to lipid peroxidation in cells. Microorganisms play an important role in maintaining plant health. In order to sustain in different environmental conditions, microorganisms induce the formation of different primary and secondary product which helps plant to alleviate different abiotic and biotic stress. ACC is known as a precursor of ethylene phytohormone. Plant growth promoting endophytes have ability to express ACC deaminase that protects plants from different abiotic and biotic stresses. It was showed by Primrose and Dilworth [93] that Mucorhiemalis produced ethylene in pure culture at a maximum rate under anaerobic condition.
4.5. 에틸렌
에틸렌은 식물에서 종자 발아, 꽃 피기, 성별 결정, 과일 숙성 및 탈락에 도움을 주는 유일한 기체 식물 호르몬입니다. 잎의 성장과 노화 조절은 에틸렌의 농도에 의존합니다. 에틸렌은 다른 식물 호르몬과 긍정적 또는 부정적으로 상호작용합니다. 아라비도프시스에서 에틸렌은 종자 휴면 상태에 대해 아브시스산과 대항적으로 작용합니다. 특정 농도를 초과하면 두 물질 모두 뿌리 연장 을 억제합니다. 에틸렌에 에틸렌 합성 억제제를 처리하면 그 농도가 감소하고 ABA가 뿌리 성장 억제에 효과적으로 작용합니다. 이는 에틸렌이 뿌리 성장 반응에서 ABA와 음의 상호작용을 한다는 것을 나타냅니다 [89]. Stepanova 등 [90]은 에틸렌이 뿌리 성장 촉진을 위해 옥신 생합성을 자극한다는 것을 발견했습니다. 그들은 WEI7 (Weak Ethylene Insensitive7)과 WEI2 전사체를 자극함으로써 트립토판 생산이 증가하며, 이는 결국 아라비도프시스 묘의 옥신 생산으로 이어진다는 것을 보여주었습니다. 에틸렌은 식물의 하부 줄기 부위에서 HOOKLESS1(N-아세틸트랜스퍼레이스 유전자 가족의 일원)을 조절함으로써 auxin 반응 인자 [ARF]의 수준을 조절하며, 이는 이러한 조직에서 잘 유지되고 정의된 차등 성장 패턴을 유발합니다 [90]. 에틸렌 농도는 초식동물과의 상호작용 후 증가하는 것으로 확인되었습니다. 이는 식물이 곤충에 의해 먹힐 때 곤충과 미생물이 분비하는 구강 분비물 때문일 수 있습니다.
에틸렌은 자연적 대사 과정과 인간의 신체 내 공기에서 발생합니다. 인간의 장내 미생물군집에는 에틸렌이 존재하며, 이는 인간에게 알려진 유전독성 발암물질(IARC 그룹 1)인 에틸렌 옥사이드를 생성할 수 있습니다. 인간의 호기 공기에서도 에틸렌이 검출되었습니다 [91]. Laurent 등 [92]은 인간에서 전신 염증 시 에틸렌이 생성된다고 보고했습니다. 전신 염증과 감염 시 활성 산소 종(ROS)은 산화 스트레스를 유발하여 불포화 지방산의 내인성 지질 과산화를 일으키고 에틸렌 형성을 초래합니다(그림 1 및 2). 호흡 폭발은 산화 스트레스의 원인이 되며, 세균 및 곰팡이 감염이 혈액 세포(단핵구 및 중성구)를 파괴하는 병원체와 만나면 발생합니다. 이 폭발로 인해 ROS가 대량으로 생성되어 산화 스트레스를 유발합니다. 그들은 Fe³⁺ 이온에 노출된 후 지방산(도코사헥사엔산과 올레산)이 산화 스트레스를 유발해 세포 내 지질 과산화를 초래하며 에틸렌을 크게 생성한다는 것을 보여주었습니다. 미생물은 식물 건강 유지에 중요한 역할을 합니다. 다양한 환경 조건에서 생존하기 위해 미생물은 식물이 다양한 비생물적 및 생물적 스트레스를 완화하는 데 도움을 주는 다양한 일차 및 이차 물질을 생성합니다. ACC는 에틸렌 식물 호르몬의 전구체로 알려져 있습니다. 식물 성장 촉진 내생균은 ACC 탈아미노효소를 발현하여 식물을 다양한 비생물적 및 생물적 스트레스로부터 보호합니다. Primrose와 Dilworth [93]은 Mucorhiemalis가 무산소 조건에서 순수 배양 시 최대 속도로 에틸렌을 생성함을 보여주었습니다.
5. Research challenges and future perspectives
The exogenous supply of these phytohormones in human body can affect the human gut microbiome and thus have an effect on metabolism and other biological activities. Many studies reported that gut microbes have the ability to produce these phytohormones but their concentration is not known. Endophytes found in fruits and vegetables having property to produce these hormones when consumed by human beings and can directly affect human health. It might show beneficial and harmful effects on human health. The genes responsible for the production of these hormones in microbes can be isolated and with the help of genetic engineering and their production can be regulated. This can be used in making drugs for treatment of various diseases, pharmaceuticals products and cosmetics. In the future research, omics data on the human whole microbiota study could be investigated to solve this unexplored mysterious question. Furthermore, the plant hormones play a key role in the signalling between plant and plant associated microbes, but its understanding between plant- animal–microbe interactions remains unexplored.
6. Concluding remark
The role of phytohormones is very well known in plants but very few studies have been done on the effect of these phytohormones in humans and other animals. Phytohormones have an effect on biological activities such as metabolic activity, disease resistance. For example, ABA and Ethylene have protective activity in metabolic diseases (Type-2 diabetes) and also induce cells’ innate immunity. Hormones like Cytokinin and auxin directly quench reactive oxygen and induce anti-oxidant defences in human body. Cytokinin can be used as an active ingredient in cosmetics such as anti-ageing cream or beauty capsules and lotions. An effective concentration of cytokinin, GA and auxin can be used in treatment of cancer and tumour.
Author's declaration
All authors are aware about this communication. This is original review article and there is no conflict of interest between the authors and others.
Author's contribution
AM contributed to the preparation of the manuscript, its formatting and collection of data for writing and preparing figures. AKG, SS, SY and SB helped in creating figures and editing the manuscript. The main idea of this manuscript was designed by AM and JPV. Final editing done by JPV and SA
Declaration of Competing Interest
All authors are aware about this communication. This is original research manuscript. There is no conflict of interest between each author and others. There is no financial conflict of interest. Funding agency properly acknowledged in manuscript.
Acknowledgement
Authors thankful to DST (DST/SEED/SCSP/STI/2020/426/G; DST/INT/SL/P-31/2021), SERB (EEQ/2021/0001083) and Banaras Hindu University-IoE (6031) for financial support for research and development for enhancing sustainable agricultural productivity and microbiome study.
References
- 1.Davies P.J. The plant hormones: their nature, occurrence, and functions. Plant Hormones. 2010:1–15. doi: 10.1007/978-1-4020-2686-7_1. [DOI] [Google Scholar]
- 2.Went F.W., Thimann K.V. Macmillan; New York: 1937. Phytohormones. [Google Scholar]
- 3.Jiang K., Asami T. Chemical regulators of plant hormones and their applications in basic research and agriculture. Biosci. Biotechnol. Biochem. 2018;82(8):1265–1300. doi: 10.1080/09168451.2018.1462693. [DOI] [PubMed] [Google Scholar]
- 4.Vildanova M.S., Savitskaya M.A., Onishchenko G.E., Smirnova E.A. The effect of plant hormones on the components of the secretory pathway in human normal and tumor cells. Cell Tissue Biol. 2014;8(5):407–415. [PubMed] [Google Scholar]
- 5.Gaspar T., Kevers C., Penel C., Greppin H., Reid D.M., Thorpe T.A. Plant hormones and plant growth regulators in plant tissue culture. In Vitro Cell. Dev. Biol. Plant. 1996;32(4):272–289. [Google Scholar]
- 6.Asami T., Nakagawa Y. Preface to the special issue: brief review of plant hormones and their utilization in agriculture. J. Pestic. Sci. 2018;43(3):154–158. doi: 10.1584/jpestics.M18-02. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Trypuc J.A., Matejczyk M., Czerpak R. N6-benzyladenine and kinetin influence antioxidative stress parameters in human skin fibroblasts. Mol. Cell. Biochem. 2016;413(1–2):97–107. doi: 10.1007/s11010-015-2642-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Blaser M.J., Cardon Z.G., Cho M.K., Dangl J.L., Donohue T.J., Green J.L., Knight R., Maxon M.E., Northen T.R., Pollard K.S., Brodie E.L. Toward a predictive understanding of Earth's microbiomes to address 21st century challenges. MBio. 2016;7(3):e00714–e00716. doi: 10.1128/mBio.00714-16. [DOI] [PMC free article] [PubMed] [Google Scholar]