Structure and function of muscle fibers and motor units - 캠브리지 대학 교과서
작성자문형철작성시간12.07.23조회수965 목록 댓글 0muscle의 흥분-수축, 이완의 최소단의 motor unit
위대한 근육생리학자 셰링턴이 제시한 개념
꼭 기억해야할 개념
1. 하나의 근육에 motor unit는 5~1500개. 평균 150개
2. 하나의 motor unit에는 1~160개의 운동종판이 있음.
3. motor unit size는 차이가 있고, 작은 motor unit부터 흥분, 수축에 동원
4. 통증, 손상, 습관에 따라 근육 동원의 패턴이 깨짐
![]() Structure and function of muscle fibers and motor units.pdf
Structure and function of muscle fibers and motor units.pdf
Introduction
The term “motor unit” was introduced by Sir Charles Sherrington, a founder of modern neurophysiology, who observed that force occurred in discrete steps when a muscle contracted in the stretch reflex [1]. He postulated that each step was produced by the all-or-none action of a single motor neuron upon the muscle fibers it innervated. Sherrington’s concept of the motor unit assumed that each muscle fiber receives innervation from only one motor neuron, and that the muscle fiber faithfully responds to every impulse of the motor neuron. These assumptions have subsequently been shown to be true in healthy adult skeletal muscles.
The motor unit has become a fundamental concept in understanding the physiology of muscle and the control of movement. A motor unit consists of one motor neuron and all the muscle fibers it innervates. The term muscle unit has been
introduced to refer to the group of muscle fibers innervated
by a given motor neuron [2]. The motor neuron and its
muscle unit are inseparable in function because each action
potential in the neuron activates all fibers of the muscle unit.
Thus motor units are the indivisible quantal elements in all
movements. The electrophysiological, metabolic, mechanical,
and anatomical properties of the motor neuron and its
muscle unit are coordinated in a manner that allows efficient
muscle contraction over a wide range of motor behaviors.
The coordinated expression of the proteins that govern these
properties reflects the interplay between the trophic control
that motor neurons exert over their muscle fibers through
activity patterns and chemical trophic factors, as well as
trophic feedback from the muscle fiber to the motor neuron.
Although most of the properties of a given motor unit
become specified during the early postnatal period of development,
physical activity and disease processes can modify
certain properties to a limited extent. In this chapter, the
basic structural and physiological properties of motor units
and muscle fibers will be introduced, with a particular
emphasis on humans and other mammals.
Anatomy of motor units
Motor neurons
Motor neurons are the only central neurons with axons that
leave the central nervous system (CNS) to innervate nonneuronal
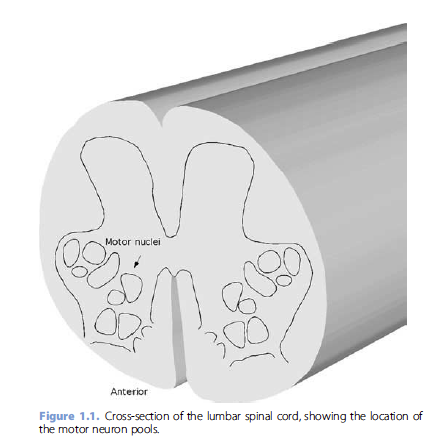
tissue. Their cell bodies are located in the anterior
horn of the gray matter of the spinal cord (Figure 1.1). The
motor neurons that innervate the same muscle cluster together
in motor nuclei that form elongated columns that generally
extend over several spinal cord segments [3]. The number of
motor neurons innervating each muscles varies, ranging from
the estimates of 30–40 motor neurons innervating the delicate
tenuissimus muscle in the cat [4] to estimates of 100–200
motor neurons innervating human thenar muscles [5, 6]. In
the lumbar and cervical enlargements of the spinal cord, the
motor neurons that innervate distal limb muscles are located
most laterally within the anterior horn, and motor neurons
innervating proximal muscles lie more medially [7, 8]. The
axons of motor neurons exit the spinal cord through the
adjacent anterior roots. When motor axons innervating the
same muscle exit from roots of several segments, they rejoin in
a muscle nerve after traversing peripheral plexuses and nerve
trunks. The muscle nerve contains motor axons innervating
the muscle and the sensory axons arising from receptors
within the muscle, such as the muscle spindles and tendon
organs.
In mammals, there are three kinds of motor neurons in the
motor nucleus. Alpha motor neurons are large cells [9, 10] that
innervate the striated muscle fibers that make up the bulk of
skeletal muscle tissue (extrafusal fibers). Gamma, or fusimotor,
neurons are considerably smaller [11] and exclusively innervate
one or more of the three types of specialized muscle fibers
within the muscle spindle – stretch receptor organs that are
present in virtually all somatic muscles [12, 13]. A third class of
motor neuron, called skeleto-fusimotor or beta motor
neurons, innervates both intra- and extrafusal muscle fibers
[14]. Beta motor neurons have been found in higher primates
[15] and probably also occur in humans. Because beta motor neurons are difficult to identify in physiological experiments,
there is little direct evidence about their properties. What
little is known indicates that the properties of beta motor
neurons and their extrafusal muscle fibers are essentially the
same as those of alpha motor neurons [16]. For this reason,
alpha and beta motor neurons will not be distinguished in this
chapter.
Alpha motor neurons have extensive dendritic trees that
receive synaptic input over their entire extent [17, 18, 19].
Their myelinated axons have large diameters with correspondingly
fast conducting velocities, ranging from 40 to 60 m/s
in human motor nerves [20]. Faster conduction velocities,
50–120 m/s, have been reported in cats and smaller mammals
[21]. The axons of motor neurons can be extremely long, up to
a meter in length for those motor neurons innervating the
distal foot muscles of a tall adult. The length and diameter
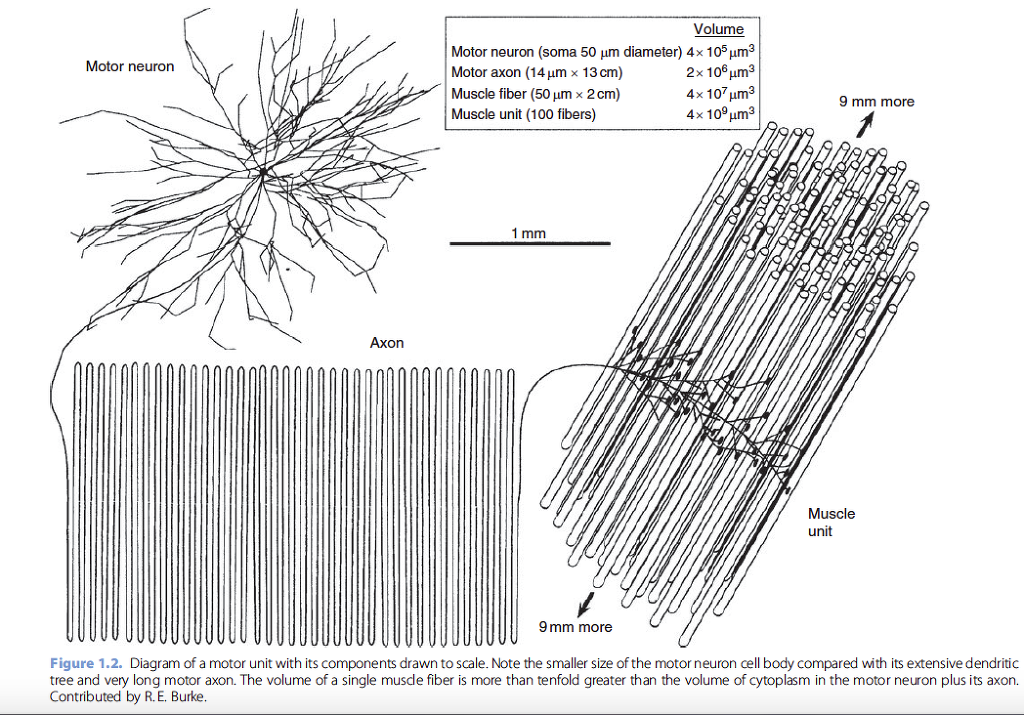
of the motor axons mean that the volume of axoplasm may
exceed the volume in the cell body and dendrites by tenfold or
more (Figure 1.2). The large metabolic demands of maintaining
the peripheral axon presumably account for the large size
of the motor neuron cell body.
Neuromuscular junctions
As the myelinated motor axons near their target muscle, they
begin to divide into tens or hundreds of terminal branches,
which lose their myelin sheaths as they near the neuromuscular
junctions (NMJs). The NMJ is a large, highly specialized
synapse between the motor nerve terminal and the muscle
fiber [22]. In somatic muscles there is only one NMJ per
muscle fiber [23], but exceptions are found in some cranial
muscles, such as the laryngeal [24] and extraocular muscles
[25]. On a given muscle fiber, the NMJ is located approximately
equidistant from its ends, allowing action potential
depolarization to spread equally to both ends from the center
of the muscle fiber. The NMJ is a complex structure that
undergoes remodeling during development and aging and in
response to denervation. At the NMJ, the motor nerve terminal
is separated from the postsynaptic muscle membrane by
a synaptic space containing basal lamina with synapse-specific
glycoproteins. On the postsynaptic side, the muscle membrane
is highly folded. Acetylcholine receptors are found on the
crests of the junctional folds apposing the vesicle release sites
on the presynaptic terminal, whereas the voltage-gated sodium
channels responsible for action potential generation are densest
in the depths of the folds [26]. NMJs exhibit structural
specializations related to the size and type of muscle fiber [27].
The structure and function of NMJs will be covered more fully
in Chapter 23.
Muscle fibers
The skeletal muscle fiber is a cylindrical, multinucleated cell
that is formed by the fusion of myoblast cells during development.
The muscle fiber has a highly organized structure, with
several distinct spatial domains. Nuclei are positioned along
the periphery of the fiber beneath the plasma membrane, or
sarcolemma. The center of the muscle fiber is packed with the
contractile apparatus, which consists of longitudinally oriented
myofibrils and scaffolding proteins. The contractile apparatus
is encircled by a network of sarcoplasmic reticulum (SR), a
form of endoplasmic reticulum specialized for calcium release
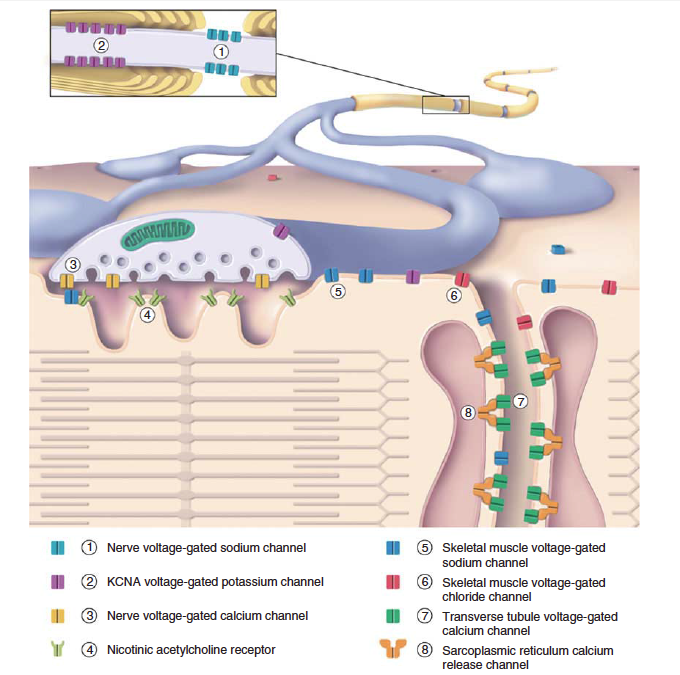
and reuptake. The sarcolemma has numerous narrow infoldings,
called T-tubules, that penetrate deep into the muscle
fiber, where they become closely apposed to regions of the
SR at specialized junctions called triads or “calcium release
junctions.” The T-tubule membrane is continuous with the
sarcolemma membrane, but it is specifically enriched in certain
membrane proteins, such as voltage-gated calcium channels,
chloride channels, and transporters (Figure 1.3) [28, 29].
The T-tubule “interior” is in continuity with the extracellular
space, although diffusion occurs more slowly from this narrow
space than at the surface membrane. The triads, where
T-tubules meet the SR, are the sites where action potential
depolarization is coupled to the mechanical contraction.
Excitation–contraction coupling occurs through protein–
protein interactions between the sarcoplasmic domains of the
voltage-gated calcium channels on T-tubule membranes and
the calcium release proteins, known as ryanodine receptors, on
the SR membrane [30].
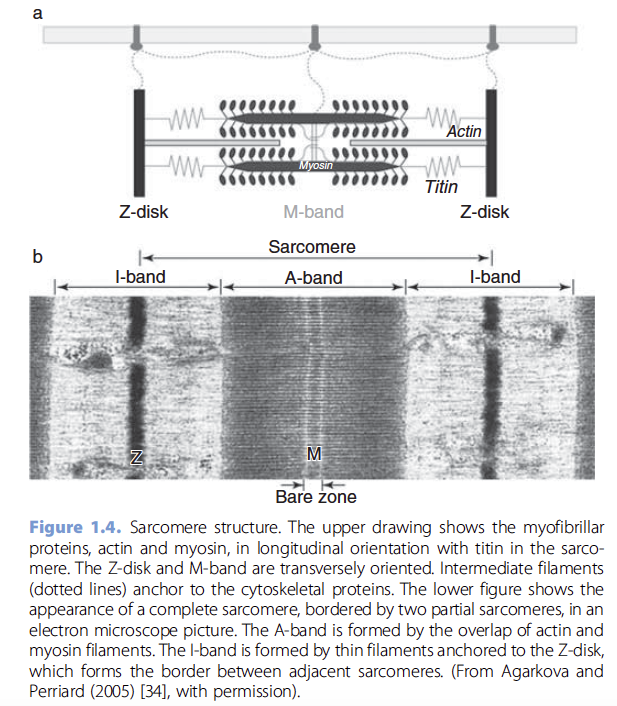
The contractile apparatus of the muscle is organized into a
series of repeated units a few microns long called sarcomeres
[31]. The sarcomere is the smallest unit of contraction. It
consists of highly organized protein assemblies that give the
muscle fiber a characteristic striated appearance (Figure 1.4b).
The sarcomere contains the myofibrils, longitudinal arrays of
thick and thin filaments that are maintained in a hexagonal
lattice by a scaffolding network (Figure 1.4a). Proteins in the
scaffolding network condense at the ends and middle of
the sarcomere to form transverse bands called Z-disks and
M-bands [32]. The thin filaments consist of filamentous actin
entwined by tropomyosin and troponin, a calcium-binding
protein. Thick filaments consist of myosin, a large molecule
with heavy and light chains. The myosin heavy chains have a
tail region and a globular head. Thick filaments are formed by
the assembly of myosin monomers with their tails centrally
and heads protruding outwards, with an antiparallel orientation
on opposite ends of the filament. Z-disks, which mark the
border between sarcomeres, serve to anchor the thin filaments.
The Z-disks are formed by an ensemble of several proteins,
including alpha-actinin. Titin, a large elastic protein spanning
from the Z-disk to the M-band, binds to the myofibrils, keeping
them centered in the sarcomere, and transmitting tension
to the Z-disk during sarcomere shortening [33]. Titin and
proteins that comprise the M-band essentially form an intrasarcomeric
cytoskeleton that maintains the regular spacing of
the thick and thin filaments [32, 34].
The myosin heads on the thick filament contain an ATPase
activity and binding sites for actin. When contraction is initiated
by a muscle fiber action potential, calcium released from
the SR binds troponin, uncovering binding sites on actin. This
leads to the formation of cross-bridges between actin and
myosin. The ATPase activity of myosin is enhanced by formation
of cross-bridges, and as ATP is hydrolyzed the crossbridge
is broken, freeing the myosin head to swivel to the next
actin-binding site. The repeated formation and cleavage of
actomyosin cross-bridges produces the sliding action of thin
and thick filaments that causes shortening of the sarcomere
and muscle contraction [35, 36]. The actomyosin cross-bridges
serve as the mechanical linkage between thick and thin filaments
for transmitting tension to the insertions of the muscle
fiber. The amount of tension is proportional to the number of
cross-bridges, reaching a maximum at sarcomere lengths when
thick and thin filaments have the greatest overlap [37, 38].
The muscle fiber has a rich cytoskeletal network underlying
the membrane and surrounding the myofibrils. In subsarcolemmal
regions, protein complexes of dystrophin, syntrophins,
and other molecules bind to F-actin and other cytoskeletal
proteins. By binding as well to intracellular domains of
membrane proteins such as sarcoglycans these effect a linkage
between the muscle interior and the extracellular matrix.
Beneath the subsarcolemmal cytoskeleton, networks of intermediate filaments, of which desmin is the most prominent,
play a role in the positioning and morphology of organelles
within the muscle (reviewed in [39, 40]). Desmin connects
Z-disks, SR, myofibrils, and other organelles to the subsarcolemmal
cytoskeleton. Mitochondria are usually found in two locations
within the muscle fiber, beneath the sarcolemma and
among the myofibrils, mostly near the Z-disks. Subsarcolemmal
and interfibrillarmitochondria appear to be functionally distinct,
with differing cytochrome content, capacity for ADP-stimulated
respiration, and susceptibility to apoptotic stimuli [41, 42].
Deficiencies of desmin lead to subsarcolemmal accumulation of
mitochondria in mice, supporting a key role for desmin in
mitochondrial positioning [43]. Intermediate filaments also bind
to proteins on the surface of lysosomes, which are relatively
sparse in normal muscle, but become prominent in some myopathies.
Glycogen particles, sometimes termed glycosomes, are
found in myofibrillar and subsarcolemmal locations.
Extracellular matrix
The muscle fiber is surrounded by an extracellular matrix
which consists of several distinct layers [44]. The innermost
layer, the basal lamina, contains the carbohydrate-rich extracellular
domains of membrane proteins, such as dystroglycan
and integrins, that interact with the muscle cytoskeleton;
secreted glycoproteins such as members of the laminin family; and a variety of ligands and proteoglycans that bind to the
extracellular matrix proteins. The outermost layer is rich in
collagen fibers, forming a connective tissue layer, the endomysium.
The extracellular matrix is specialized at the NMJ, containing
synaptic laminins, ligands such as agrin, and the
enzyme acetylcholinesterase. The basal lamina and the extracellular
matrix molecules play a key role in supporting muscle
fiber development and regeneration after injury. Lying beneath
the basal lamina are satellite cells, myogenic precursors that are
able to proliferate and differentiate into myoblasts [45].
Muscles
Most mammalian muscle fibers are only a few centimeters
long, much shorter than the length of most muscles. The
length of a muscle fiber is thought to be limited by the need
for sarcomeres to be activated nearly simultaneously to produce
an effective contraction, which in turn is limited by the
time needed for an action potential to travel the length of
the muscle fiber. The conduction velocity of muscle fibers is
relatively slow, in the range of 2–10 m/s [46, 47]. To achieve an
effective mechanical action over a larger length, groups of
muscle fibers, called fascicles, are bound together by perimysial
connective tissue to form a muscle. Muscle fascicles are
arranged in various ways that allow a common direction of
force to be delivered to the muscle’s points of origin and
insertion [48]. There are two general schemes [49]: pinnate,
in which the muscle fibers are oriented at an angle to the
muscle’s primary direction of force; and parallel, in which
the orientation of muscle fibers is the same as the force vector.
In pinnate muscles, the fascicles are arranged in parallel
bundles, often in a feather-like pattern along one or more
tendinous aponeuroses (Figure 1.5a). Muscles with pinnate
architecture have relatively limited distensibility, but can
deliver large output forces. Pinnation is commonly seen in
muscles with relatively short lever arms that operate over a
limited range of physiological lengths, for example the gastrocnemius
muscles of the leg. At the other extreme are muscles
with parallel arrangements of interdigitated muscle fascicles,
staggered at different longitudinal locations along a web-like
intramuscular stroma (Figure 1.5b; [47, 50]). This arrangement
allows a small amount of slippage of fascicles past each
other, and is commonly seen in muscles that span multiple
joints or undergo large changes in length during movement.
As might be expected, some muscles exhibit mixtures of these
designs (e.g., tibialis anterior in the cat; [51]). A few long,
strap-like muscles, such as the biceps femoris, have two or
more bellies arranged in series separated by tendinous inscriptions
that create distinct anatomical compartments [52].
Most muscles have an optimal range of working lengths.
When muscles are stretched during natural movements, they
offer some resistance. Most of the tension is related to the
number of cross-bridges between overlapping thick and thin
filaments [37, 38]. Additional contributions from tendons and
internal connective tissue enter into consideration primarily
when a muscle is stretched beyond its optimal working range.
Because connective tissue is less elastic than muscle fibers,
tension rises quickly at these lengths. Contributions from
connective tissue to muscle length–tension curves are referred
to as passive, in contrast to the active contributions from
the myofibrillar cross-bridges. Passive contributions to muscle
tension differ between healthy and diseased muscle.
Degenerative muscle diseases, or even the prolonged disuse of
muscles, such as after a stroke, may result in markedly
increased connective tissue within the muscle with stiffness
and increased resistance to stretch [53].
Functional organization of motor units
Distribution of motor unit fibers
The spatial distribution of muscle fibers belonging to an individual
motor unit has been studied experimentally with the
glycogen depletion technique [54]. In this method, prolonged
stimulation of a motor axon is used to deplete muscle fibers of
endogenous glycogen stores, enabling the depleted fibers to be
identified histochemically. The glycogen depletion method
showed that muscle fibers belonging to the same motor unit
were arranged in a mosaic fashion among muscle fibers
belonging to other motor units [54, 55]. Relatively few muscle
fibers from the same unit occurred immediately adjacent to
one another [56, 57]. Statistical studies suggest that the distribution
of fibers in single units is basically random [58]. Nevertheless,
the arrangement of the muscle unit’s fibers must
accommodate to the internal architecture of the parent muscle
to produce a meaningful pattern of force. In pinnate muscles,
fibers from one motor unit were found to be scattered more or
less evenly through territories that were relatively large, but
smaller than the total cross-section of the muscle (Figure 1.6).
In multicompartment muscles, motor unit fibers were usually
distributed only within one compartment [59]. However, there
are examples, such as the extensor digitorum muscle of the monkey forelimb, in which fibers of one motor unit are
distributed among several compartments to exert a common
force on multiple tendons [60].
Electromyographic (EMG) studies of single motor units in
humans suggest a similar spatial organization of muscle unit
fibers. Using a technique called scanning EMG, in which a
motor unit action potential is recorded as an electrode is
advanced in successive steps of 50 μm through the muscle,
Stalberg and colleagues [61, 62] recorded territories with
cross-sectional areas of 2–10mm for single motor units in
the biceps and tibialis anterior muscles. Within the same
region of muscle, they found that several dozen motor units
had overlapping territories. For an individual motor unit, at
some places the muscle fiber action potentials were grouped,
and separated from other regions, suggestive of fractions of the
muscle unit innervated by different branches of the motor
axon (arrows, Figure 1.7).
One way to describe the size of a motor unit is according to
its innervation ratio: the number of muscle fibers innervated
by a given motor neuron. The number of muscle fibers innervated by one motor neuron varies widely between different
muscles. In humans, innervation ratios have been estimated
by dividing an estimate of the total number of muscle
fibers in a muscle by counts of the number of large axons in
cross-sections of the muscle nerve. Such calculations have
produced estimates of innervation ratios ranging from less
than a dozen for the extraocular muscles to over a thousand
for motor units of large limb muscles (Table 1.1) [63, 64].
Physiological methods have also been used to estimate the
number of motor units innervating certain muscles, and these
studies have also shown similar ranges [6]. However, using the
glycogen depletion method to identify the fibers of individual
motor units in animals, Burke and Tsairis [56] found considerable
variation in the innervation ratios for different units
within a given muscle. The innervation ratio of the motor unit
is a major factor governing its force output. Variation in
innervation ratios is likely to provide much of the variability
in force output produced by different motor units within a
muscle [65, 66].
Muscle fiber types
For more than a century, it has been recognized that mammalian
muscles fall into two general groups: dark “red” muscles
with slow contraction times and lighter “white” muscles with
fast contraction times. Histological and physiological studies
have shown that most muscles contain a mixture of muscle
fibers with differing contraction speeds and force outputs;
muscles composed of purely fast or slow muscle fibers are
exceptional (for reviews see [67, 68]). The isoform of the
myosin heavy chain (MHC) expressed in the muscle fiber is
one of the most important factors influencing the speed of
contraction, because the rate of ATP hydrolysis determines the
speed of cross-bridge cycling and sarcomere shortening [69,
70]. Other factors affecting the contractile speed of muscle
fibers include the isoforms of the calcium reuptake and release
proteins expressed and the density of the SR [71, 72, 73, 74].
There are three major isoforms of MHC expressed in adult
human limb muscles: MHC I, also called slow myosin; and the
two fast isoforms, MHC IIA and MHC IIX (also called MHC
IID). Subtypes of these isoforms, as well as embryonic and
neonatal forms of MHCs, generate further diversity. The fast
and slow isoforms of myosin were first able to be distinguished
histochemically because of their differing amounts of ATPase
activity at acid and alkaline pH [75]. This histochemical difference
allowed fast and slow muscle fibers to be classified into
two types. Fast and slow muscle fiber types are further subdivided
by their dependence on aerobic or anaerobic metabolic
pathways. Muscle fibers that utilize oxidative metabolism for
energy needs have abundant mitochondria and lipid droplets.
In contrast, muscle fibers using anaerobic pathways for energy
tend to be richer in glycolytic enzymes with more abundant
glycogen stores. Histochemical methods for demonstrating
mitochondrial enzymes combined with myosin ATPase activity
have traditionally been used to define three major types of
muscle fiber in adult human limb muscles, described below.
The histochemical properties of different fiber types correspond
fairly well to their contractile properties, allowing muscle
fibers to be grouped into a small number of types by either
histochemical or physiological measures. It should be recognized,
however, that qualitative and quantitative differences in
expression of fiber-type-specific proteins generate a continuous
range of physiological properties.
Type 1 muscle fibers have a slow twitch and use oxidative
metabolism. Type 1 fibers express MHC I, the slow isoform of
myosin, and contain many mitochondria. These muscle fibers
can be visualized histochemically by strong myosin ATPase
activity at low pH and by dense staining for mitochondrial
enzymes such as NADH dehydrogenase (i.e., nicotinamide
adenine dinucleotide, reduced) and SDH (i.e., succinate dehydrogenase)
(Table 1.2). Compared to Type 2 fibers, their SR is
less abundant, and it contains a slower isoform of the SR
calcium ATPase. Type 1 fibers contain myoglobin, a protein
that binds oxygen and confers a red color, and have a rich
capillary blood supply [76]. The metabolic profile and vascularization
render Type 1 muscle fibers highly resistant to
fatigue, and thus suitable for sustained contraction under
aerobic conditions. The acronym “SO,” slow oxidative, is used
by some to denote these fibers.
Type 2 muscle fibers are fast-twitch fibers, expressing fast
isoforms of myosin which exhibit strong ATPase activity at
alkaline pH. There are several subtypes of Type 2 fibers, but
two major subtypes occur in human limb muscles. Type 2A
fibers express the MHC IIA isoform of myosin. Compared to
Type 1 fibers, their SR is denser, and expresses isoforms of
calcium handling proteins that allow a more rapid cycling of
calcium ions from SR [71, 72, 73]. Mitochondria are relatively
abundant in Type 2A fibers. In addition Type 2A fibers contain glycolytic enzymes, such as phosphorylase, and have
abundant glycogen stores. These metabolic properties allow
Type 2A to function under aerobic and anaerobic conditions,
and provide them with a fairly high resistance to fatigue. Type
2A fibers have been denoted by the acronym “FOG” because
they are fast twitch with oxidative and glycolytic metabolic
capabilities. The third major muscle fiber type that occurs in
human limb muscles is the Type 2B fiber. Type 2B fibers
express the fastest isoform of myosin, MHC IIX (also known
as IID). Their SR is dense and contains a fast isoform of SR
calcium ATPase. Type 2B fibers have relatively sparse mitochondria,
but contain glycolytic enzymes and stores of glycogen.
Type 2B muscle fibers fatigue easily, but are suitable for
short bursts of anaerobic exercise. The acronym “FG,” fast,
glycolytic, is sometimes used.
Other isoforms of myosin are found in specialized muscle
or at different developmental stages. In a number of animal
species, Type 2B fibers express a very fast form of myosin, the
MHC IIB isoform, particularly in muscles with very fast speeds
of contraction [77, 78, 79, 80]. In humans, MHC IIB expression
has been reported in some cranial muscles [81] but it is
not expressed to a significant extent in limb muscles. Immature
forms of myosin are expressed by muscle fibers prior to
completing their differentiation during development [82, 83].
Fibers expressing immature forms of myosin that stain for
ATPase activity at acid and alkaline pH, Type 2C fibers, are found
in small numbers in normal adult limb muscles. The Type 2C
profile occurs in regenerating fibers, which can be common in
several muscular dystrophies. Muscle spindles also express a
mixture of immature and slow isoforms of myosin [84].
The classification of the major muscle fiber types by their
pattern of MHC expression agrees relatively well with the
histochemical classification of fiber Types 1, 2A, and 2B that
is based on myosin ATPase activity at differing pH. However,
histochemical methods are relatively insensitive to hybrid
muscle fibers expressing more than one MHC isoform. Hybrid
muscle fibers can be demonstrated with immunocytochemical
methods or in-situ hybridization for different isoforms of
MHCs [80, 85]. Combinations of MHC IIA with IIx expression
are relatively common in Type 2 fibers, for example [85,
86]. In some muscles hybrid fibers make up a sizeable fraction
of the muscle fibers [78, 79, 85]. Hybrid fibers may play a role
in the ability of muscle fibers to undergo rapid adaptations in
response to training and use [87, 88, 89, 90].
Association of motor unit types with muscle
fiber types
All muscle fibers belonging to the same motor unit have the
same type, as judged from their staining for ATPase activity
[54, 91, 92] and MHC isoforms [93, 94, 95]. Within a muscle
unit the fibers also appear to have similar metabolic enzyme
capacities [94, 96]. It is, therefore, assumed that muscle fibers
within the motor unit also have essentially identical mechanical
properties. Edstrom and Kugelberg [54] were the first to
use the glycogen depletion method to examine the association
between the mechanical properties and histochemical characteristics
of the muscle fibers of individual motor units for two
types of fast-twitch motor unit in rats. Burke and coworkers
[55, 56] later used the same approach to examine the histochemistry
of muscle fibers within the full range of physiologically
identified motor units in the cat gastrocnemius muscle. In
these studies, motor neurons were characterized physiologically
with intracellular recordings, including stimulation with
short stimulus trains while measuring force output and prolonged stimulation to deplete glycogen stores in active
muscle fibers. Burke and coworkers found that motor units
differed in several mechanical properties, not just the speed of
contraction. These properties included the magnitudes of force
produced by individual twitches (twitch force) and the maximal
force produced by repetitive stimulation (tetanic force),
resistance to fatigue during sustained activation, and the ratio
of the twitch to the tetanic force [67]. These properties each
exhibited continuous distributions that initially made it problematic
to define distinct groups of motor units. However, two
criteria were found that permitted relatively clear clustering of
motor units into fast and slow groups in the cat: a “fatigue
index” based on the decline in force output during a defined
sequence of intermittent tetanization and a “sag property”
based on the shape of unfused isometric tetanic contractions
(Figure 1.8) [55, 91, 92, 97, 98]. Using these criteria, Burke and
colleagues were able to define three main types of motor units:
Type FF (fast twitch, fatigable), Type FR (fast twitch, fatigue
resistant) and Type S (slow twitch, fatigue resistant). Some
fast-twitch units exhibited fatigue resistance intermediate
between those of FF and FR units and were, therefore, referred
to as F(int) or FI [56, 92, 99]. Physiologically, there was a
perfect match between S, FR and FF motor units with the
histochemically defined muscle fiber Types 1, 2A, and 2B,
respectively (Table 1.2; see also [97, 98, 100]). They also found
some evidence that fibers in the minority F(int) unit type were
histochemically distinct from the three main types [56, 98].
These same physiological criteria have been used with somewhat
more variable success in classifying motor units in rat
muscles (e.g., [101, 102]). It is possible that some of the
variation in properties such as contraction time within a given
motor unit type are associated with hybrid combinations of
myosin isoforms, but this remains to be investigated
systematically.
Motor units in human muscles
There is a wealth of information available from EMG studies
in humans about the behavior of motor units in normal and
diseased muscle, and it has been known for some time that
fast- and slow-twitch muscle fibers coexist in human muscle
[103]. However, for obvious technical reasons, it is difficult to
examine the mechanical responses of individual motor units
under the controlled conditions possible in animal experiments.
Denny-Brown and Pennybacker [104] were the first
to record individual twitches from the fasciculations of motor
units in patients with motor neuron disease, using an indirect
pneumatic transducer. Buchthal and Schmalbruch [105] used a
mechanical transducer attached to a needle inserted into
tendons, plus intramuscular stimulation of small nerve
branches, to demonstrate that small groups of human motor
units in normal muscles generate a wide range of twitch
speeds, which varied in relation to the predominant local fiber
type (see also [106]). The introduction of spike-triggered computer
averaging into clinical neurophysiology made it possible
to record the responses of individual motor units with greater
assurance [107]. In this technique, discharges of single motor
units during steady voluntary contractions are used to trigger
an averaging computer while measuring the force produced by
an appendage (e.g., a finger) attached to a force transducer.
There are two limitations of this technique. First, the recorded
twitch responses are not isolated twitches but rather components
of unfused tetani, leading to errors in estimating the
twitch forces and contraction times [108, 109]. Intra-neural
stimulation of single motor axons to produce twitches has
been used in an attempt to overcome this problem [110, 111,
112]. Secondly, the mechanical responses measured can be
significantly degraded by the compliance of components
between the active muscle fibers and the force transducer,
including tendons of various lengths. Despite these technical
limitations, most of the contractile properties measured from
human motor units are generally similar to those from animals
[113, 114]. There is disagreement about whether fatigability
and “sag” can be used to classify human motor unit types in
the same manner as in animals, and whether force measurements
relate to the fatigability in the same way [110, 112, 115].
However, when motor units have been identified by glycogen
depletion in muscle biopsy samples, these properties were
consistent with histochemical identification [116]. Overall,
the available physiological evidence and correspondence with the histochemical classification strongly suggest that the basic
characteristics of Types S, FF, and FR human motor units are
similar to those described for the cat and rat.
Functional correlates of fiber properties
and motor unit types
It is clear that many factors contribute to mechanical properties
of the different motor unit types: in addition to the expression
of MHC isoforms, there are fiber-type-specific differences
in myosin light chains, troponin and tropomyosin proteins,
proteins involved in calcium release and reuptake, and sarcotubular
structures [72, 73, 74, 117]. It seems likely that the
“sag” property, which differs sharply in fast and slow units, is
produced by interactions among these factors [67, 68, 118].
Resistance to fatigue is directly related to the oxidative capacity
of the different fiber types (Table 1.2; [91, 119]), as well as to
their mitochondrial content [72] and local capillary supply
[120]. These correlations are certainly causally related. The
forces produced by individual motor units can vary by over
two orders of magnitude during tetanization, and this
variation is correlated with motor unit type (Figure 1.8 and
Table 1.2). The force produced by a motor unit is a function of
the effective cross-sectional area of its muscle fibers and the
specific force output of that fiber type per unit area. Estimation
of the effective cross-sectional area must take into account the
effective innervation ratio [121], which may approximate the
actual innervation ratio in pinnate muscles [91] but would be
less in interdigitated muscles which have unit fibers in serial
arrays (Figure 1.5). In general, Type 1 and 2A fibers have
smaller diameters than Type 2B, making fiber area an important
component of the equation. In humans, Type 2 fibers
exhibit the greatest variability in diameter; in general fiber
diameters tend to be larger in men than women [122]. There
is some controversy about whether specific force output, which
cannot be measured directly, differs between units with Type 1
and 2 muscle fibers [65, 69, 91, 92].
Motor neurons and synaptic specializations
In view of the differences between muscle fiber types, it is not
surprising that the motor neurons that innervate them exhibit
corresponding physiological differences (Table 1.3; reviewed
by [67]). In general, motor neurons of Type S motor units have
slower axonal conduction velocities, longer durations of postspike
hyperpolarized after-potentials (AHPs), and higher
whole-cell input resistance values than the cells that innervate
either FR or FF motor units. The AHP duration is particularly
important because it is a key factor that controls the rate of
motor neuron firing; motor neurons of Type S units have the
longest AHPs and generally fire more slowly than those of FR
or FF units. When examined with intracellular labeling
methods, the motor neurons of Type S units tend to be smaller
in membrane area than Type FF cells; Type FR motor neurons
are intermediate in size [9, 10]. There is no systematic difference between axonal conduction velocities of FF and FR
unit groups [123]. Although the distributions of motor neuron
properties are continuous and exhibit large overlaps when
sorted according to muscle unit type, the relative excitability
of the motor neurons to depolarizing currents injected directly,
measured as the rheobase (the amount of current
required to produce action potentials reliably), is more closely
related to unit type than other measures [124, 125]. The
rheobase data imply that intrinsic motor neuron excitability
varies according to the sequence S>FR>FF, which has
important implications for the recruitment order of motor
units (Figure 1.9).
The strength of several synaptic inputs to motor neurons
shows type-related differences that are undoubtedly related to
the way in which the various types of motor units are used
during activity. For example, the average amplitudes of monosynaptic
excitatory postsynaptic potentials (EPSPs) produced
in motor neurons by group Ia muscle spindle afferents, which
are largely responsible for the stretch reflex, are ordered as
S>FR>FF (Table 1.2) [126, 127]. The same ordering is
evident with the disynaptic inhibition produced by stimulation
of group Ia afferents from antagonist muscles [126] and with
disynaptic recurrent inhibition produced by Renshaw interneurons
activated from motor axon collaterals [128]. The
organization of synaptic efficacy is a key factor that controls
the function of motor unit populations [129], and for most
inputs to motor neurons, the ordering of synaptic efficacy
follows the size principle. However, there is evidence that
certain cutaneous inputs and supraspinal systems, notably
the rubrospinal tract, tend to excite relatively high-threshold
motor neurons while inhibiting low-threshold cells [130, 131,
132], a pattern opposite to that found in group Ia excitation.
Although there would be potential advantages to competing
control systems that could bypass low-threshold, slow-twitch
motor units that are slow to relax, the idea that large, ...
더 없음.