신경과학은 알아두면 정말 유용할때가 많다.
인체가 느끼는 모든 감각의 상행로에 대한 탐구
의학을 하는 사람에게는 필수적 지식이다.
panic bird...
![]() Ascending Sensory Pathways.pdf
Ascending Sensory Pathways.pdf
A variety of sensory receptors scattered throughout the body can become activated by exteroceptive, interoceptive, or proprioceptive input. Exteroceptive input relays sensory information about the body’s interaction with the external environment. Interoceptive input relays information about the body’s internal state, whereas proprioceptive input conveys information about position sense from the body and its component parts.
Each receptor is specialized to detect mechanical, chemical, nociceptive (L. nocere, “to injure,” “painful”), or thermal stimuli. Activation of a sensory receptor is converted into nerve impulses and this sensory input is then conveyed via the fibers of the cranial or spinal nerves to their respective relay nuclei in the central nervous system (CNS). The sensory information is then further processed as it progresses, via the ascending sensory systems (pathways), to the cerebral cortex or to the cerebellum. Sensory information is also relayed to other parts of the CNS where it may function to elicit a reflex response, or may be integrated into pattern-generating circuitry.
The ascending sensory pathways are classified according to the functional components (modalities) they carry as well as by their anatomical localization. The two functional categories are the general somatic afferent (GSA) system, which transmits sensory information such as touch, pressure, vibration, pain, temperature, stretch, and position sense from somatic structures; and the general visceral afferent (GVA) system,which transmits sensory information such as pressure, pain, and other visceral sensation from visceral structures.
- 일반체성구심감각 시스템은 촉각, 압각, 진동각, 통증, 온도, 늘어남, 위치감각 을 전달함.
- 일반내장구심감각 시스템은 압각, 통증 그리고 다른 내장감각을 전달함.
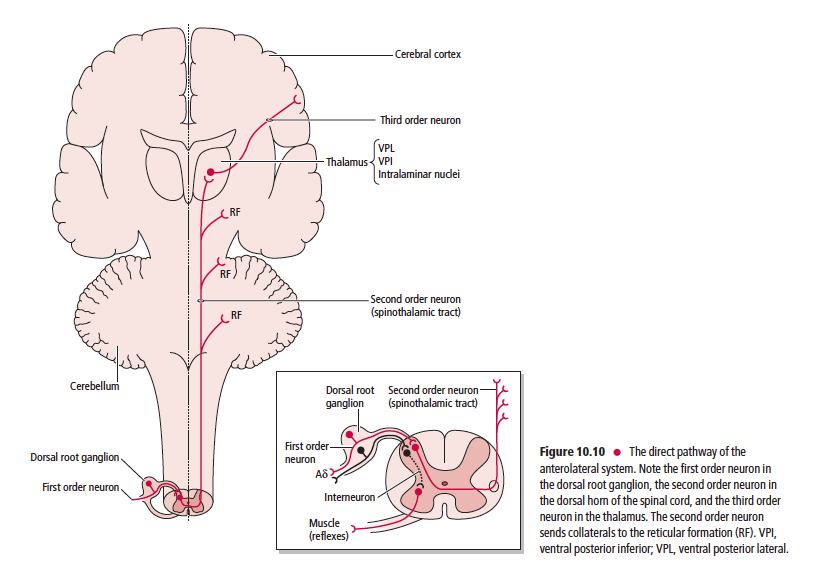
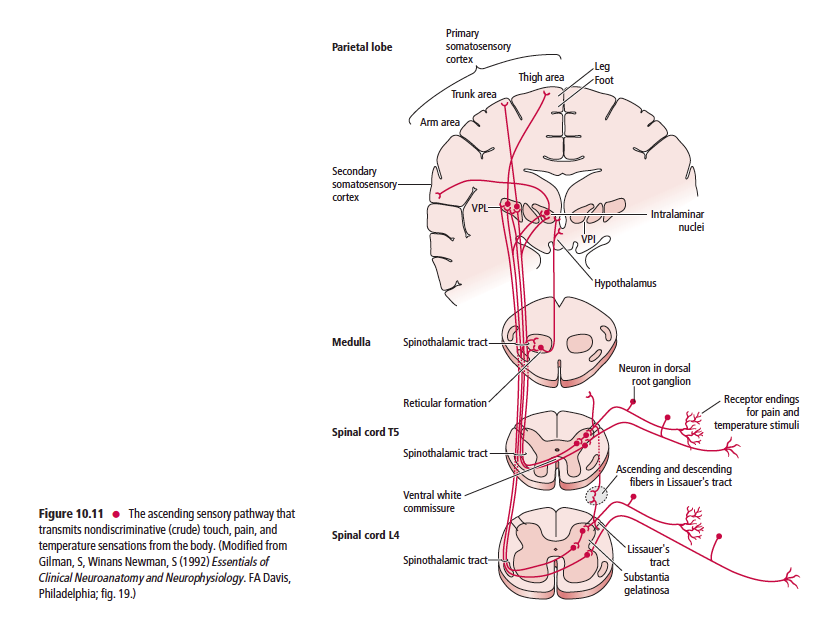
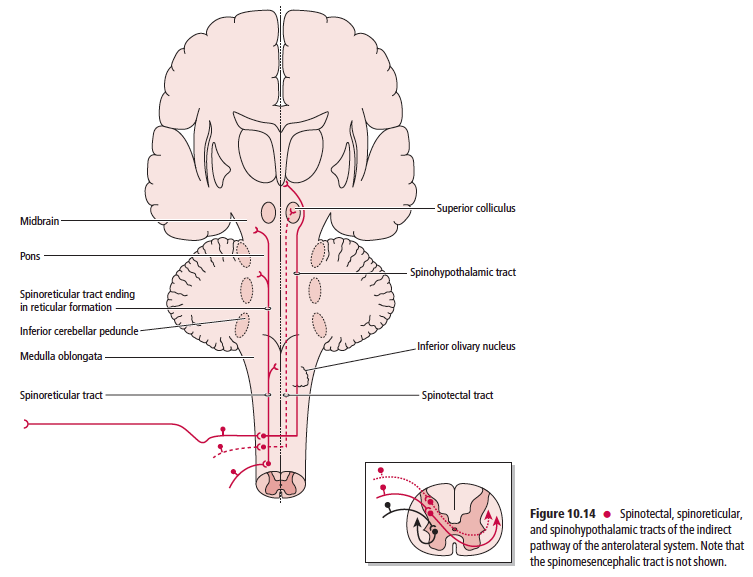
Anatomically, the ascending sensory systems consist of three distinct pathways: the anterolateral system (ALS), the dorsal column–medial lemniscal (DCML) pathway, and the somatosensory pathways to the cerebellum. The anterolateral system, which includes the spinothalamic, spinoreticular, spinomesencephalic, spinotectal, and spinohypothalamic tracts, relays predominantly pain and temperature sensation, as well as nondiscriminative (crude or poorly localized) touch, pressure, and some proprioceptive sensation (Table 10.1).
- 상행로는 세가지. ALS, DCML, somatosensory to the cerebellum 경로가 있음.
- ALS 경로는 spinothalmic, spinoreticular, spinomesencephalic, spinotectal, spinohypothalamic가 있어 통증, 온도, 촉각, 온도각, 압력, 약간의 고유수용감각을 전달함.
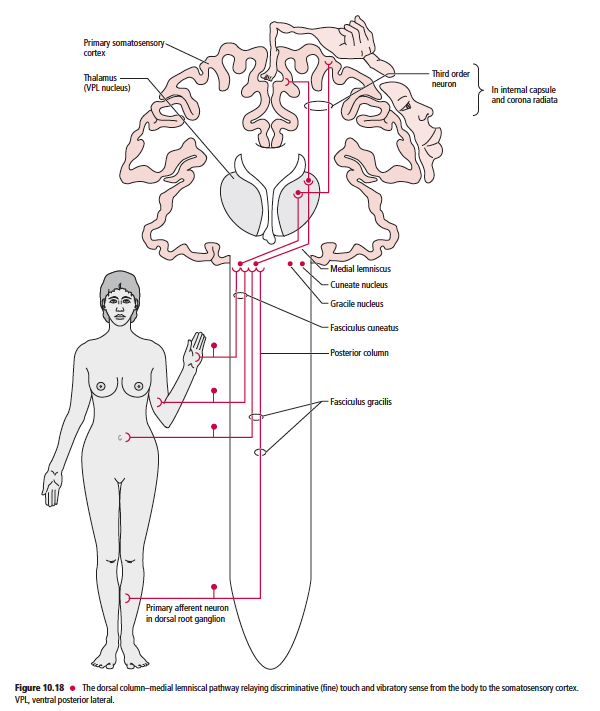
The dorsal column–medial lemniscal pathway (which includes the fasciculus gracilis, fasciculus cuneatus, and medial lemniscus) relays discriminative (fine) tactile sense, vibratory sense, and position sense (Table 10.1). The somatosensory pathways to the cerebellum, which include the anterior, posterior, and rostral spinocerebellar, as well as the cuneocerebellar tracts, relay primarily proprioceptive (but also some pain and pressure) information (Table 10.1).
- DCML경로는 fasciculus gracilis, fasciculus cuneatus가 있어서 분별감각, 진동각, 위치감각 등을 전달함.
The ascending sensory pathways are the main avenues by which information concerning the body’s interaction with the external environment, its internal condition, and the position and movement of its parts, reach the brain. One similarity shared by all three ascending sensory pathways from the body (not including the head or face) is that the first order neuron cell bodies reside in the dorsal root ganglia. It is interesting to note that conscious perception of sensory information from external stimuli is mediated by the spinothalamic and DCML pathways to the ventral posterior lateral nucleus of the thalamus, whereas sensations that do not reach consciousness are mediated by the spinoreticular, spinomesencephalic, spinotectal, spinohypothalamic, and the anterior, posterior, and rostral spinocerebellar, and cuneocerebellar tracts. These tracts terminate in the reticular formation, mesencephalon, hypothalamus and cerebellum, respectively.
Sensory input may ultimately elicit a reflex or other motor response because of the functional integration of the ascending (somatosensory) pathways, the cerebellum, and the somatosensory cortex, as well as the motor cortex and descending (motor) pathways. Furthermore, descending projections from the somatosensory cortex, as well as from the raphe nucleus magnus and the dorsolateral pontine reticular formation to the somatosensory relay nuclei of the brainstem and spinal cord, modulate the transmission of incoming sensory impulses to higher brain centers.
This chapter includes a description of the sensory receptors and the ascending sensory pathways from the body, whereas the ascending sensory pathways from the head, transmitted mostly by the trigeminal system, are described in Chapter 15.
SENSORY RECEPTORS
Although sensory receptors vary according to their morphology, the velocity of conduction, and the modality to
which they respond, as well as to their location in the body, they generally all function in a similar fashion. The stimulus to which a specific receptor responds causes an alteration in the ionic permeability of the nerve endings, generating a receptor potential that results in the formation of action
potentials. This transformation of the stimulus into an electrical
signal is referred to as sensory transduction.
Some receptors that respond quickly and maximally at
the onset of the stimulus, but stop responding even if the
stimulus continues, are known as rapidly adapting (phasic)
receptors. These are essential in responding to changes but
they ignore ongoing processes, such as when one wears a
wristwatch and ignores the continuous pressure on the skin
of the wrist. However, there are other receptors, slowly
adapting (tonic) receptors, that continue to respond as long
as the stimulus is present.
Sensory receptors are classified according to the source
of the stimulus or according to the modality to which they respond. It is important to note that, in general, receptors do
not transmit only one specific sensation.
Classification according to stimulus source
Receptors that are classified according to the source of the
stimulus are placed in one of the following three categories:
exteroceptors, proprioceptors, or interoceptors.
Exteroceptors are close to the body surface and are specialized to detect sensory information from the external environment
1 Exteroceptors are close to the body surface and are specialized
to detect sensory information from the external
environment (such as visual, olfactory, gustatory, auditory,
and tactile stimuli). Receptors in this class are sensitive
to touch (light stimulation of the skin surface),
pressure (stimulation of receptors in the deep layers of the
skin, or deeper parts of the body), temperature, pain, and
vibration. Exteroceptors are further classified as teloreceptors
or contact receptors:
• teloreceptors (G. tele, “distant”), include receptors
that respond to distant stimuli (such as light or
sound), and do not require direct physical contact
with the stimulus in order to be stimulated;
• contact receptors, which transmit tactile, pressure,
pain, or thermal stimuli, require direct contact of the
stimulus with the body.
Proprioceptors transmit sensory information from muscles, tendons, and
joints about the position of a body part, such as a limb in space
2 Proprioceptors transmit sensory information from
muscles, tendons, and joints about the position of a body
part, such as a limb in space. There is a static position
sense relating to a stationary position and a kinesthetic
sense (G. kinesis, “movement”), relating to the movement
of a body part. The receptors of the vestibular system
located in the inner ear, relaying sensory information
about the movement and orientation of the head, are also
classified as proprioceptors.
Interoceptors detect sensory information concerning the status of the body’s
internal environment
3 Interoceptors detect sensory information concerning the
status of the body’s internal environment, such as stretch,
blood pressure, pH, oxygen or carbon dioxide concentration,
and osmolarity.
Classification according to modality
Receptors are further classified into the following three
categories according to the modality to which they respond:
nociceptors, thermoreceptors, and mechanoreceptors(Table 10.2).
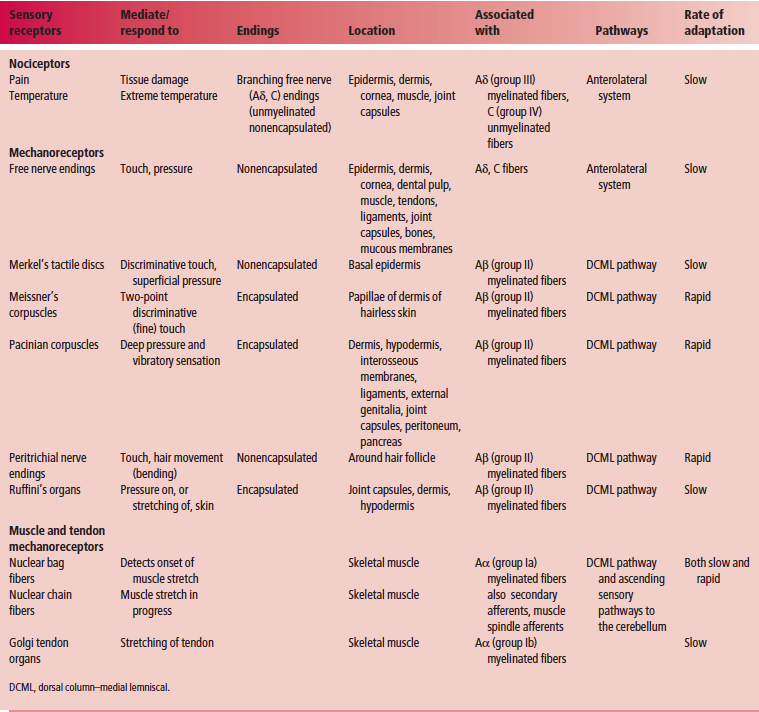
Nociceptors
Nociceptors are rapidly adapting receptors that are sensitive to noxious or
painful stimuli
Nociceptors are rapidly adapting receptors that are sensitive
to noxious or painful stimuli. They are located at the peripheral
terminations of lightly myelinated free nerve endings
of type Aδ fibers, or unmyelinated type C fibers, transmitting
pain. Nociceptors are further classified into three types.
1 Mechanosensitive nociceptors (of Aδ fibers), which are
sensitive to intense mechanical stimulation (such as
pinching with pliers) or injury to tissues.
2 Temperature-sensitive (thermosensitive) nociceptors
(of Aδ fibers), which are sensitive to intense heat and cold.
3 Polymodal nociceptors (of C fibers), which are sensitive
to noxious stimuli that are mechanical, thermal, or chemical
in nature. Although most nociceptors are sensitive
to one particular type of painful stimulus, some may
respond to two or more types.
Nociception is the reception of noxious sensory information
elicited by tissue injury, which is transmitted to the CNS
by nociceptors. Pain is the perception of discomfort or an
agonizing sensation of variable magnitude, evoked by the
stimulation of sensory nerve endings.
Thermoreceptors
Thermoreceptors are sensitive to warmth or cold
Thermoreceptors are sensitive to warmth or cold. These
slowly adapting receptors are further classified into three
types.
1 Cold receptors, which consist of free nerve endings of
lightly myelinated Aδ fibers.
2 Warmth receptors, which consist of the free nerve
endings of unmyelinated C fibers that respond to increases
in temperature.
3 Temperature-sensitive nociceptors that are sensitive to
excessive heat or cold.
Mechanoreceptors
Mechanoreceptors are activated following physical deformation of the skin,
muscles, tendons, ligaments, and joint capsules in which they reside
Mechanoreceptors, which comprise both exteroceptors and
proprioceptors, are activated following physical deformation
due to touch, pressure, stretch, or vibration of the skin,
muscles, tendons, ligaments, and joint capsules, in which they
reside. A mechanoreceptor may be classified as nonencapsulated
or encapsulated depending on whether a structural
device encloses its peripheral nerve ending component.
Nonencapsulated mechanoreceptors
Nonencapsulated mechanoreceptors are slowly adapting and include free
nerve endings and tactile receptors
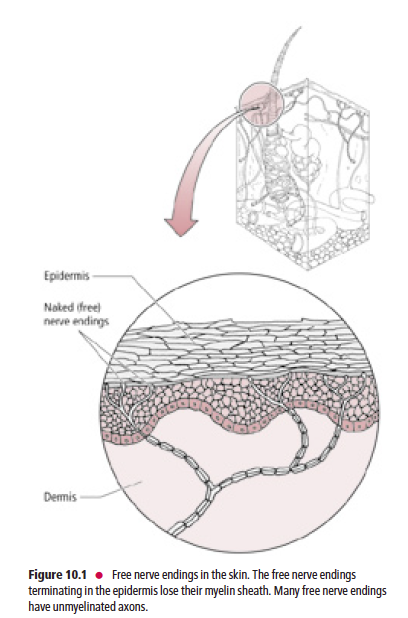
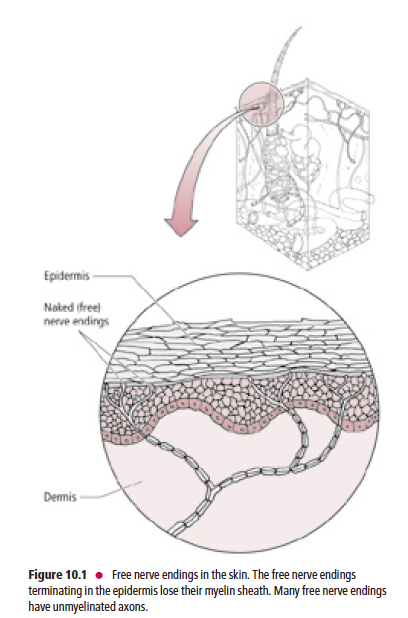
Free nerve endings (Fig. 10.1) are present in the epidermis, dermis, cornea, dental pulp, mucous membranes of the oral and nasal cavities and of the respiratory, gastrointestinal, and urinary tracts, muscles, tendons, ligaments, joint capsules, and bones. The peripheral nerve terminals of the free nerve endings lack Schwann cells and myelin sheaths. They are stimulated by touch, pressure, thermal, or painful stimuli.
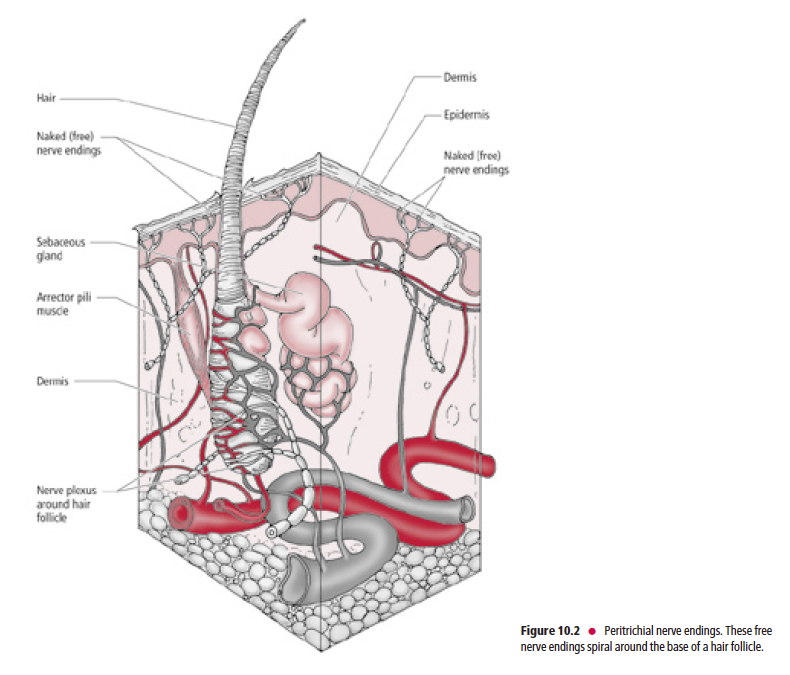
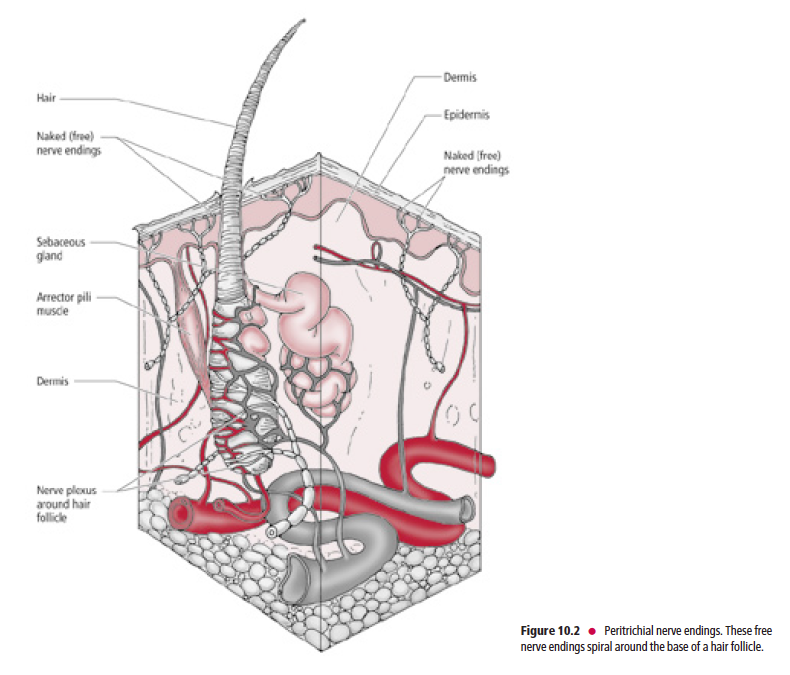
Peritrichial nerve endings (Fig. 10.2) are specialized members of this category. They are large-diameter, myelinated,
Aβ fibers that coil around a hair follicle below its associated sebaceous gland. This type of receptor is stimulated only
when a hair is being bent.
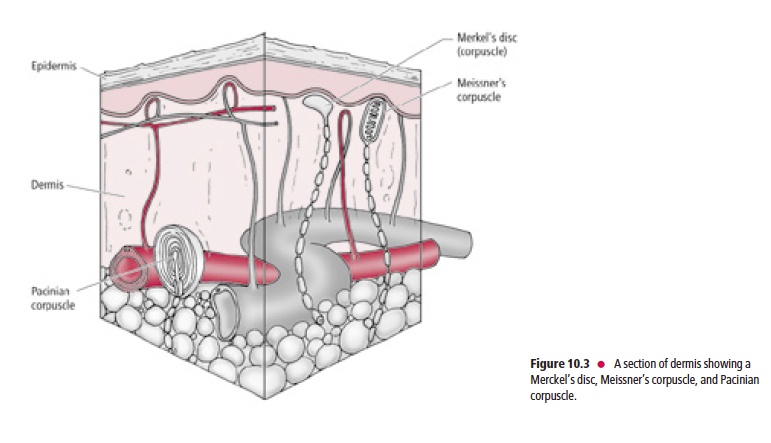
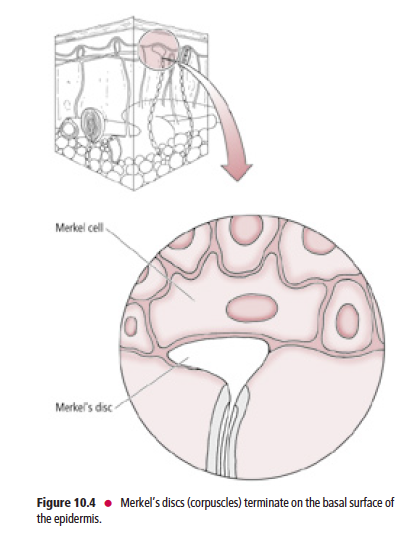
Tactile receptors (Fig. 10.3) consist of disc-shaped, peripheral nerve endings of large-diameter, myelinated, Aβ fibers.
Each disc-shaped terminal is associated with a specialized epithelial cell, the Merkel cell, located in the stratum basale
of the epidermis.
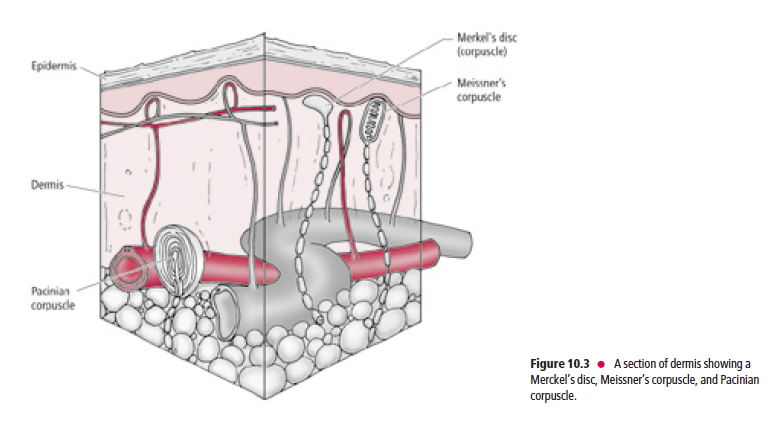
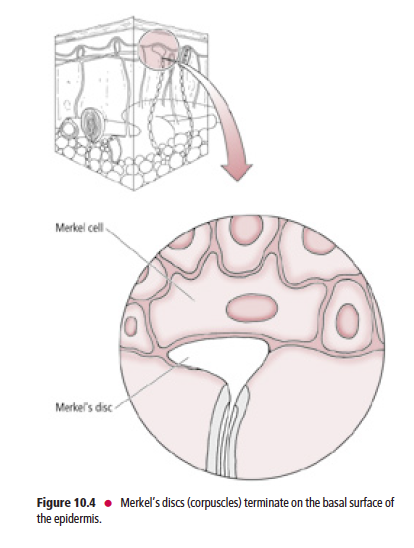
These receptors, frequently referred to as Merkel’s discs (Fig. 10.4), are present mostly in glabrous (hairless), and occasionally in hairy skin. Merkel’s discs respond to discriminative touch stimuli that facilitate the distinguishing of texture, shape, and edges of objects.
Encapsulated mechanoreceptors
Encapsulated mechanoreceptors include Meissner’s corpuscles, pacinian corpuscles, and Ruffini’s end organs.
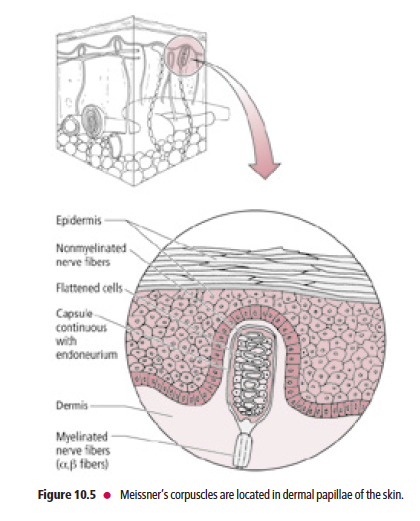
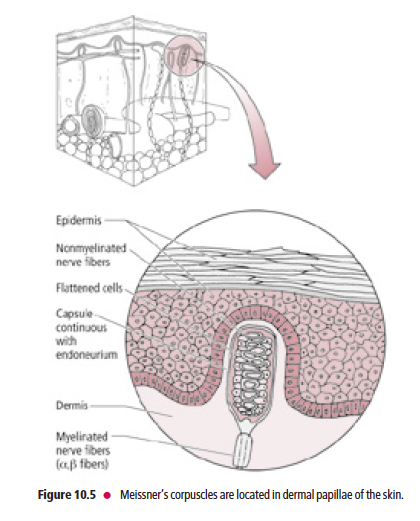
Meissner’s corpuscles are present in the dermal papillae of glabrous skin of the lips, forearm, palm, and sole, and in the connective tissue papillae of the tongue
Meissner’s corpuscles (Fig. 10.5) are present in the dermal papillae of glabrous skin of the lips, forearm, palm, and sole, as well as in the connective tissue papillae of the tongue. These corpuscles consist of the peripheral terminals of Aβ
fibers, which are encapsulated by a peanut-shaped structural device consisting of a stack of concentric Schwann cells
surrounded by a connective tissue capsule. They are rapidly adapting and are sensitive to two-point tactile (fine) discrimination, and are thus of great importance to the visually impaired by permitting them to be able to read Braille.
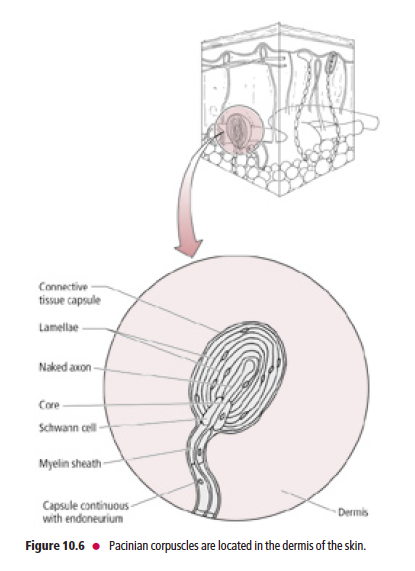
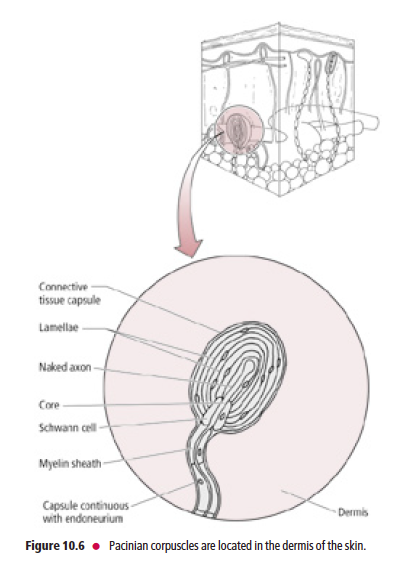
Pacinian corpuscles are the largest of the mechanoreceptors
Pacinian corpuscles (Fig. 10.6), the largest of the mechanoreceptors, are rapidly adapting and resemble an
onion in cross-section. Each Pacinian corpuscle consists of Aβ-fiber terminals encapsulated by layers of modified
fibroblasts that are enclosed in a connective tissue capsule. Pacinian corpuscles are located in the dermis, hypodermis,
interosseous membranes, ligaments, external genitalia, joint capsules, and peritoneum, as well as in the pancreas. They are more rapidly adapting than Meissner’s corpuscles and are believed to respond to pressure and vibratory stimuli,
including tickling sensations.
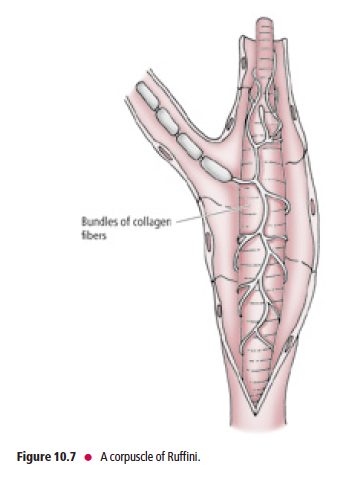
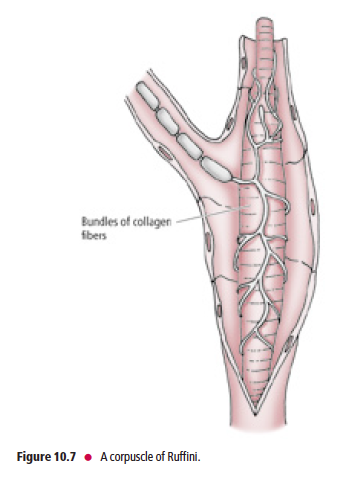
Ruffini’s end organs are located in the joint capsules, dermis, and underlying
hypodermis of hairy skin
Ruffini’s end organs (corpuscles of Ruffini) (Fig. 10.7) are located in joint capsules, the dermis, and the underlying
hypodermis of hairy skin. The unmyelinated peripheral terminals of Aβ myelinated fibers are slowly adapting. They
intertwine around the core of collagen fibers, which is surrounded by a lamellated cellular capsule. Ruffini’s end organs respond to stretching of the collagen bundles in the skin or joint capsules and may provide proprioceptive information. Muscle spindles and Golgi tendon organs (GTOs) are also encapsulated mechanoreceptors, but, due to their specialized function, they are discussed separately.
Muscle spindles and Golgi tendon organs
The mucle spindles and GTOs detect sensory input from the skeletal muscle and transmit it to the spinal cord
where it plays an important role in reflex activity and motor control involving the cerebellum. In addition, sensory input from these muscle receptors is also relayed to the cerebral cortex by way of the DCML pathway, which mediates information concerning posture, position sense, as well as movement and orientation of the body and its parts.
Muscle spindles
Structure and function
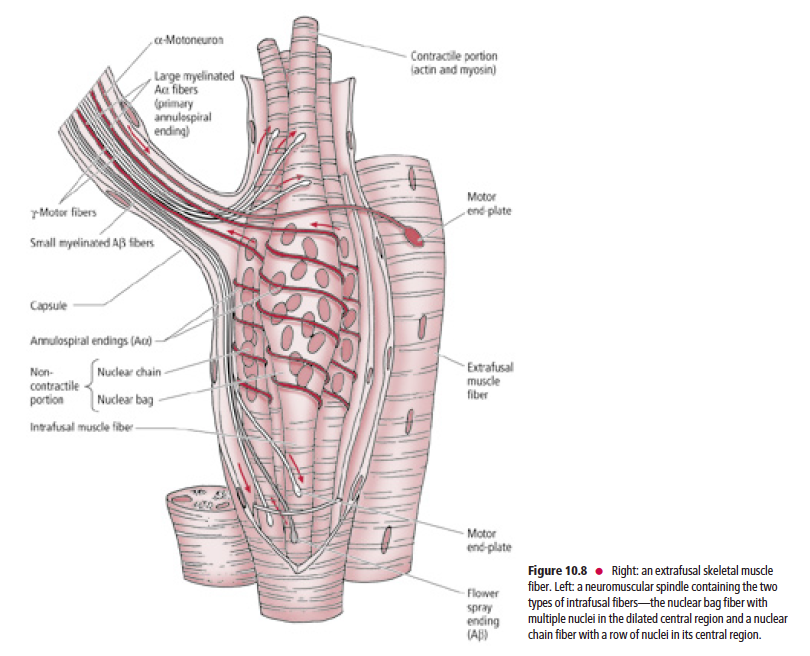
Skeletal muscle consists of extrafusal and intrafusal fibers
Extrafusal fibers are ordinary skeletal muscle cells constituting
the majority of gross muscle, and their stimulation results
in muscle contraction. Muscle spindles, composed of small
bundles of encapsulated intrafusal fibers, are dispersed
throughout gross muscle. These are dynamic stretch receptors
that continuously check for changes in muscle length.
Each muscle spindle is composed of two to 12 intrafusal
fibers enclosed in a slender capsule, which in turn is surrounded by an outer fusiform connective tissue capsule
whose tapered ends are attached to the connective tissue
sheath surrounding the extrafusal muscle fibers (Fig. 10.8).
The compartment between the inner and outer capsules contains
a glycosaminoglycan-rich viscous fluid.
There are two types of intrafusal fibers based on their
morphological characteristics: nuclear bag fibers and nuclear
chain fibers. Both nuclear bag and nuclear chain fibers possess
a central, noncontractile region housing multiple nuclei,
and a skeletal muscle (myofibril-containing) contractile portion
at each end of the central region. The nuclear bag fibers
are larger, and their multiple nuclei are clustered in the “baglike”
dilated central region of the fiber. The nuclear chain
fibers are smaller and consist of multiple nuclei arranged
sequentially, as in a “chain” of pearls, in the central region of
the fiber.
Each intrafusal fiber of a muscle spindle receives sensory innervation via the
peripheral processes of pseudounipolar sensory neurons
Each intrafusal fiber of a muscle spindle receives sensory
innervation via the peripheral processes of pseudounipolar
sensory neurons whose cell bodies are housed in dorsal root
ganglia, or in the sensory ganglia of the cranial nerves (and in
the case of the trigeminal nerve, within its mesencephalic
nucleus). Since the large-diameter Aα fibers spiral around the
noncontractile region of the intrafusal fibers, they are known
as annulospiral or primary endings. These endings become
activated at the beginning of muscle stretch or tension. In
addition to the annulospiral endings, the intrafusal fibers,
mainly the nuclear chain fibers, also receive smaller diameter,
Aβ peripheral processes of pseudounipolar neurons. These
nerve fibers terminate on both sides of the annulospiral ending,
are referred to as secondary or flower spray endings,
and are activated during the time that the stretch is in
progress (Fig. 10.8).
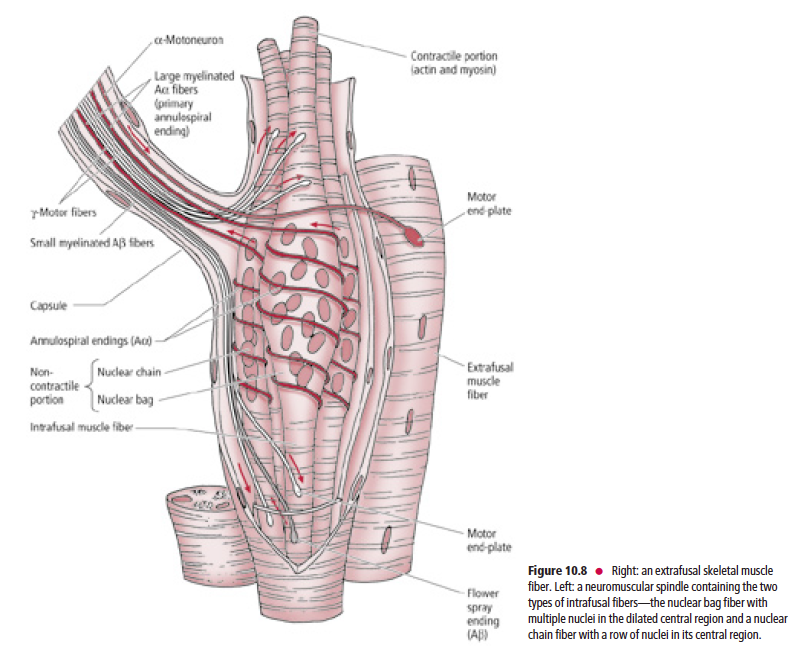
In addition to sensory innervation, intrafusal fibers also receive motor
innervation via gamma motoneurons that innervate the contractile portions
of the intrafusal fibers, causing them to contract In addition to the sensory innervation, intrafusal fibers
also receive motor innervation via gamma motoneurons
(fusimotor neurons) that innervate the contractile portions of
the intrafusal fibers, causing them to undergo contraction.
Since the intrafusal fibers are oriented parallel to the longitudinal
axis of the extrafusal fibers, when a muscle is stretched,
the central, noncontractile region of the intrafusal fibers is
also stretched, distorting and stimulating the sensory nerve
endings coiled around them, causing the nerve endings to
fire. However, when the muscle contracts, tension on the
central noncontractile region of the intrafusal fibers decreases
(which reduces the rate of firing of the sensory nerve endings
coiled around it).
During voluntary muscle activity simultaneous stimulation
of the extrafusal fibers by the alpha motoneurons, and
the contractile portions of the intrafusal fibers by the gamma
motoneurons, serves to modulate the sensitivity of the
intrafusal fibers. That is, the gamma motoneurons cause
corresponding contraction of the contractile portions of the
intrafusal fibers, which stretch the central noncontractile
region of the intrafusal fibers. Thus, the sensitivity of the
intrafusal fibers is constantly maintained by continuously
readapting to the most current status of muscle length. In this
fashion the muscle spindles can detect a change in muscle
length (resulting from stretch or contraction) irrespective of
muscle length at the onset of muscle activity. It should be
noted that even though they contract, the intrafusal fibers,
due to their small number and size, do not contribute to
any significant extent to the overall contraction of a gross
muscle.
Simple stretch reflex
The simple stretch reflex, whose mechanism is based on the
role of the intrafusal fibers, functions to maintain muscle
length caused by external disturbances. As a muscle is
stretched, the intrafusal fibers of the muscle spindles are
also stretched. This in turn stimulates the sensory afferent
annulospiral and flower spray endings to transmit this
information to those alpha motoneurons of the CNS (spinal
cord, or cranial nerve motor nuclei) that innervate the agonist
(stretched) muscle as well as to those motoneurons that
innervate the antagonist muscle(s). The degree of stretching
is proportional to (or related to) the load placed on the
muscle. The larger the load, the more strongly the spindles
are depolarized and the more extrafusal muscle fibers are in
turn activated. As these alpha motoneurons of the stretched
muscle fire, they stimulate the contraction of the required
number of extrafusal muscle fibers of the agonist muscle.
The alpha motoneurons of the antagonist muscle(s) are
inhibited so the antagonist muscle relaxes. The simple reflex
arc involves the firing of only two neurons—an afferent
sensory neuron and an efferent motoneuron—providing
dynamic information concerning the changes of the load
on the muscle and position of the body region in threedimensional
space.
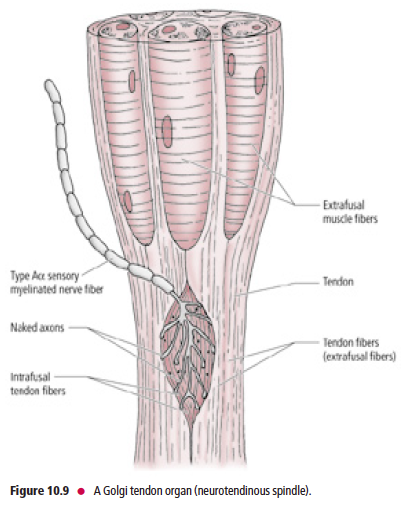
Golgi tendon organs
GTOs (neurotendinous spindles) are fusiform-shaped receptors located at
sites where muscle fibers insert into tendons
Unlike muscle spindles, which are oriented parallel to the longitudinal axis of the extrafusal muscle fibers, GTOs are in series. Furthermore, GTOs do not receive motor innervation as the muscle spindles do. GTOs consist of interlacing intrafusal collagen bundles enclosed in a connective tissue capsule (Fig. 10.9).
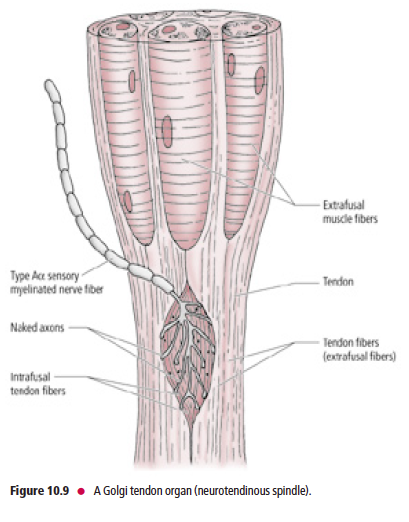
A large-diameter, type Aα sensory fiber,
whose cell body is housed in a dorsal root sensory ganglion
or a cranial nerve sensory ganglion, passes through the
capsule and then branches into numerous delicate terminals
that are interposed among the intrafusal collagen bundles.
The central processes of these Aα afferent neurons enter
the spinal cord via the dorsal roots of the spinal nerves to
terminate and establish synaptic contacts with inhibitory
interneurons that, in turn, synapse with alpha motoneurons
supplying the contracted agonist muscle.
Combined muscle spindle and Golgi tendon organ
functions during changes in muscle length
During slight stretching of a relaxed muscle, the muscle spindles are
stimulated while the GTOs remain undisturbed and quiescent; with further
stretching both the muscle spindles and GTOs are stimulated During muscle contraction, as the muscle shortens, tension
is produced in the tendons anchoring that muscle to bone,
compressing the nerve fiber terminals interposed among the
inelastic intrafusal collagen fibers. This compression activates
the sensory terminals in the GTOs, which transmit this
sensory information to the CNS, providing proprioceptive
information concerning muscle activity and preventing the
placement of excessive forces on the muscle and tendon. In
contrast, the noncontractile portions of the muscle spindles
are not stretched, and are consequently undisturbed. The
contractile regions of the muscle spindles, however, undergo
corresponding contraction that enables them to detect a
future change in muscle length (resulting from stretch or
contraction).
During slight stretching of a relaxed muscle, the muscle
spindles are stimulated whereas the GTOs remain undisturbed
and quiescent. During further stretching of the
muscle, which produces tension on the tendons, both
the muscle spindles and the GTOs are stimulated. Thus
GTOs monitor and check the amount of tension exerted on
the muscle (regardless of whether it is tension generated by
muscle stretch or contraction), whereas muscle spindles
check muscle fiber length and rate of change of muscle length
(during muscle stretch or contraction).
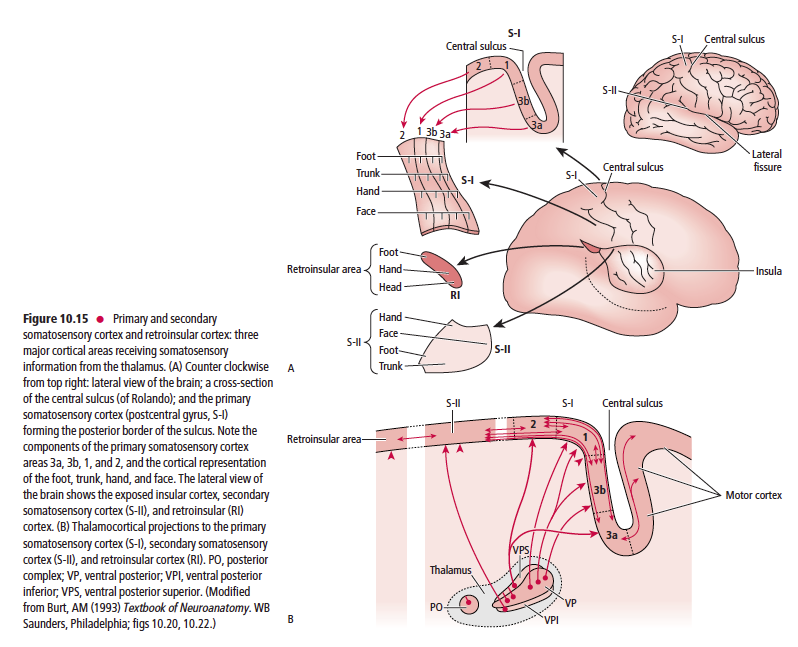
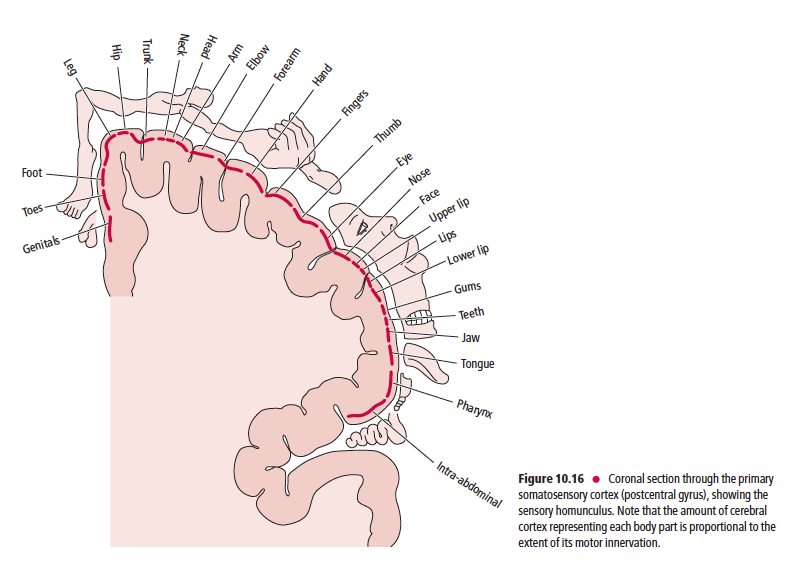
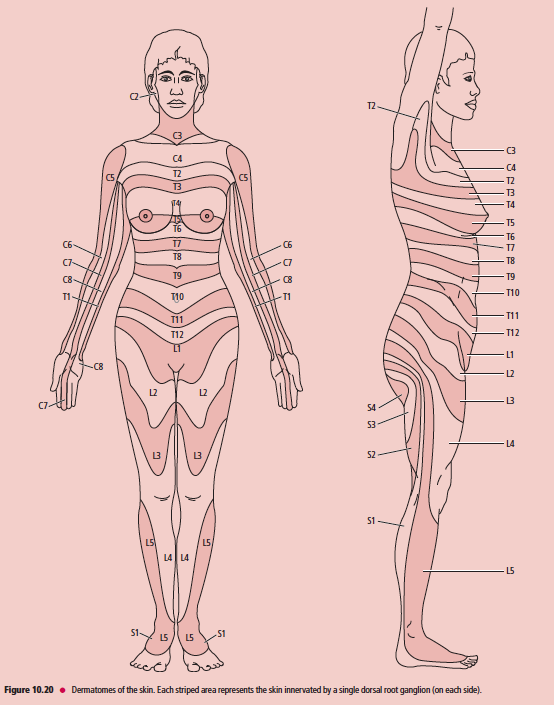
이후 중추신경전달은 그림만 올림.