beyond reason
간단히 정리하면 히스톤 아세틸화는 히스톤아세틸트랜스퍼라제(HATs)와 히스톤 디아세틸라제(HDACs) 2개의 효소에 의해 결정되는데 .. 암과같은 질환은 HDACs가 억제되지 못한 것임. 그래서 HDACs억제제는 암세포를 사멸시키는 작용을 함.
포도씨에 많은 레스베라트롤은 HDACs 억제제로 알려져 임상중임

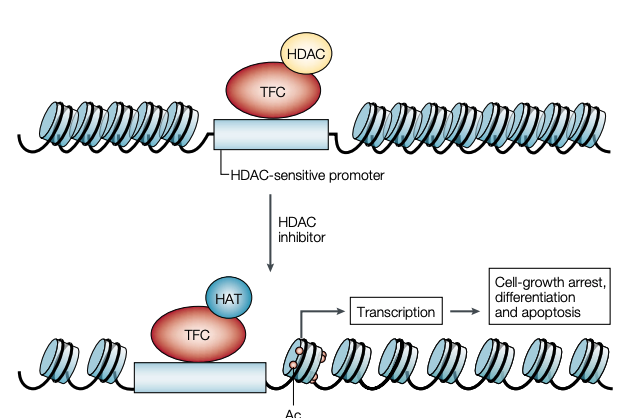
Figure 2 | Proposed mechanism of action of histone deacetylase inhibitors. With inhibition of histone deacetylases (HDACs) by HDAC inhibitors such as suberoylanilide hydroxamic acid, histones are acetylated, and the DNA that is tightly wrapped around a deacetylated histone core relaxes. We propose that there are specific sites in the promoter region of a subset of genes (for example, SP1 sites) that recruit the transcription factor complex (TFC) with HDAC and that the accumulation of acetylated histones in nucleosomes leads to increased transcription of this subset of genes (for example, CDKN1A, which encodes WAF1), which, in turn, leads to downstream effects that result in cell-growth arrest, differentiation and/or apoptotic cell death and, as a consequence, inhibition of tumour growth. Ac, acetyl group; HAT, histone acetyltranferase.
- HDACs 억제기전(HDACs가 억제된다는 의미는 암세포의 전사가 바뀌어 암세포 사멸의 길을 간다는 의미)
- suberoylanilide hydroxamic acid와 같은 HDAC억제제는 히스톤이 아세틸화되고 DNA는 단단하게 탈아세틸화된 히스톤 core왁스로 둘러싸이게 함.

Histone deacetylases and cancer: causes and therapies
- Paul A. Marks,
- Richard A. Rifkind,
- Victoria M. Richon,
- Ronald Breslow,
- Thomas Miller &
- William K. Kelly
Nature Reviews Cancer volume 1, pages194–202(2001)Cite this article
1488 Accesses
1523 Citations
9 Altmetric
Key Points
Histone acetylation is determined by the activities of two classes of enzyme: histone acetyltransferases (HATs) and histone deacetylases (HDACs). - 히스톤 아세틸화의 활성은 히스톤아세틸트랜스퍼라제(HATs)와 히스톤 디아세틸라제(HDACs) 2개의 효소에 의해 결정됨.
Histones are the core protein components of nucleosomes and their acetylation status regulates, in part, gene expression. - 히스톤은 뉴클레오좀의 core 단백질이고 히스톤의 아세틸화 상태는 유전자 발현을 조절함.
Deacetylated histones are generally associated with silencing gene expression; so the acetylation of histones is generally associated with derepression of gene expression. - 탈아세틸화된 히스톤은 silencing 유전자발현과 관계가 있음. 그래서 히스톤의 아세틸화는 일반적으로 유전자 발현의 탈억제와 관련됨.
Aberrant acetylation is associated with several solid tumours and haematological malignancies. - 비정상적인 아세틸화는 몇가지 고형암과 혈액암의 발생과 연관되어 있음.
Several types of compound have been identified that inhibit HDACs and cause accumulation of acetylated histones in normal and tumour tissues. They also inhibit transformed cell growth in vitro and in vivo. - 정상과 암조직에서 몇가지 물질이 HDACs을 억제하고 아세틸화된 히스톤의 축적을 일으키는 것을 연구로 확인되었음.
Inhibition of HDACs with hydroxamic-acid-based hybrid polar compounds — for example, suberoylanilide hydroxamic acid (SAHA) — alters transcription of very few expressed genes. - HDACs의 억제는 유전자발현의 전사를 조절함.
Several HDAC inhibitors are in clinical trials with cancer patients. They are well tolerated, cause accumulation of acetylated histones in peripheral mononuclear cells and tumours and, more importantly, have clinical activity with objective tumour regression. - 몇가지 HDAC 억제제는 암환자에게 사용되고 있음. 그것은 말초 mononuclear cell과 암조직에서 내성이 잘 생기고 아세틸화된 히스톤의 축적을 일으킴. 더 중요한 것은 암세포 소멸효과가 있음.
Abstract
Together, histone acetyltransferases and histone deacetylases (HDACs) determine the acetylation status of histones. This acetylation affects the regulation of gene expression, and inhibitors of HDACs have been found to cause growth arrest, differentiation and/or apoptosis of many tumours cells by altering the transcription of a small number of genes. HDAC inhibitors are proving to be an exciting therapeutic approach to cancer, but how do they exert this effect?
Neoplastic transformation — characterized by inappropriate cell proliferation and/or altered patterns of cell death — does not necessarily destroy the potential that cells have to differentiate and/or apoptose under appropriate environmental conditions1 . Various chemical agents can induce these processes in transformed cells2–5, and this is being exploited by cancer researchers as they attempt to develop anticancer therapies using these agents. Among the most effective of these are histone deacetylase (HDAC) inhibitors4,5. Histone proteins organize DNA into nucleosomes, which are regular repeating structures of chromatin. The acetylation status of histones alters chromatin structure, which, in turn, is involved in gene expression6,7. Two classes of enzyme can affect the acetylation of histones — histone acetyltransferases (HATs) and HDACs6–9. Altered HAT or HDAC activity has been identified in several cancers10–13. A number of HDAC inhibitors have been characterized that inhibit tumour growth in vitro and in vivo at amounts that have little or no toxicity14–26, and several of these are in clinical trials. These compounds act very selectively to alter the transcription of fewer than 2% of expressed genes27.
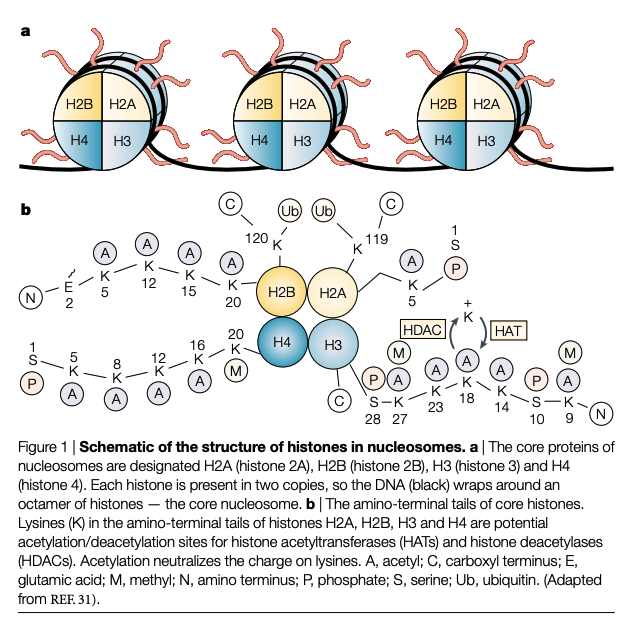
Nucleosome structure
Nucleosomes, which represent the principal protein–nucleic-acid relationship that is found in chromatin, are involved in the regulation of gene expression. The nucleosome contains 146 base pairs of DNA wrapped around the core histone octamer. The histone octamer is composed of two copies of each of H2A, H2B, H3 and H4. These proteins are very basic and highly conserved throughout evolution. All four core histones have an amino-terminal tail. This is lysine rich and contains about half of the positively charged residues and most of the post-translational modification sites of the core histones (FIG. 1). However, the functions of these post-translational modifications are largely unclear29–31. Histone H1 is also important in determining the level of DNA condensation, but as it is not regulated by acetylation, it will not be discussed further. The N-terminal tail of the histones passes through and around the enveloping DNA double helix. The modification of the structure of these N-terminal tails of histones — by acetylation/deacetylation — is crucial in modulating gene expression, as it affects the interaction of DNA with transcription-regulatory nonnucleosomal protein complexes32–34 (FIG. 2). In light of the importance of the N-terminal lysinerich histone tails with regard to DNA-regulatory protein interactions, it has been proposed that a regulatory ‘code’ resides in the pattern of post-translational modifications (such as phosphorylation, acetylation, methylation and/or ADP-ribosylation), of which these tails are the target32 (FIG. 1). This ‘code’ is read by the non-histone proteins and multiprotein complexes that form the transcription-activating and transcription-repressing molecular machinery. Of these modifications, histone acetylation has received the most analysis33.