좋은 책으로 근수축에 대하여 완벽하게 이해해보자
사랑, 존경, 공존
기쁨, 감사, 축복
chapter 8. 골격근의 구조와 기능
인체는 600개 이상의 수의적 골격근을 지니고 있는데, 이는 전체 체중의 40-50%를 차지함. 새로운 연구 증거는 근섬유가 내분비기관이고, 신체의 다양한 기관계 조정에 중요한 역할을 한다고 제시함.
골격근의 구조
근육세포 자체, 신경조직, 혈관, 결합조직을 포함하는 것이 골격근임.
근막, 근외막, 근주막, 근섬유속, 근내막
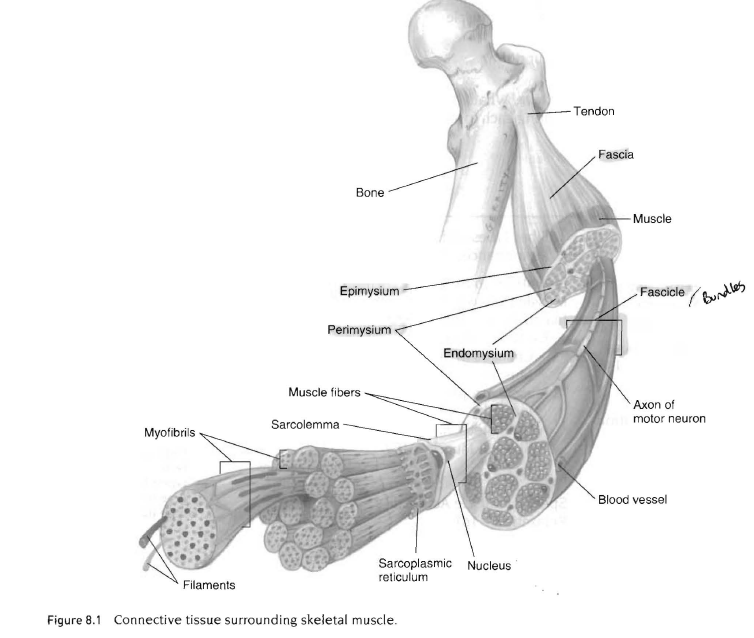
근섬유의 독특한 형태에도 불구하고 근세포는 다른세포와 마찬가지로 같은 조직들을 가짐. 즉 미토콘드리아, 리소좀 등을 가지고 있음. 그러나 다른 세포와 달리 많은 핵을 가지고 있음.
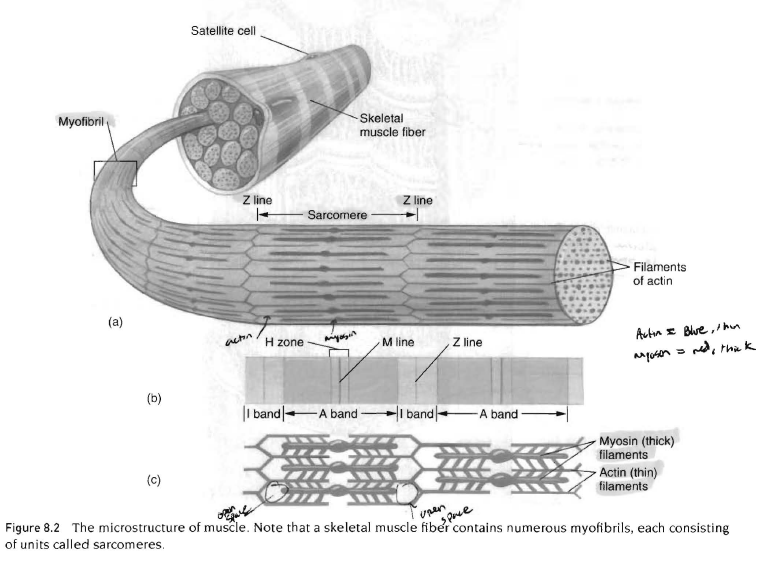
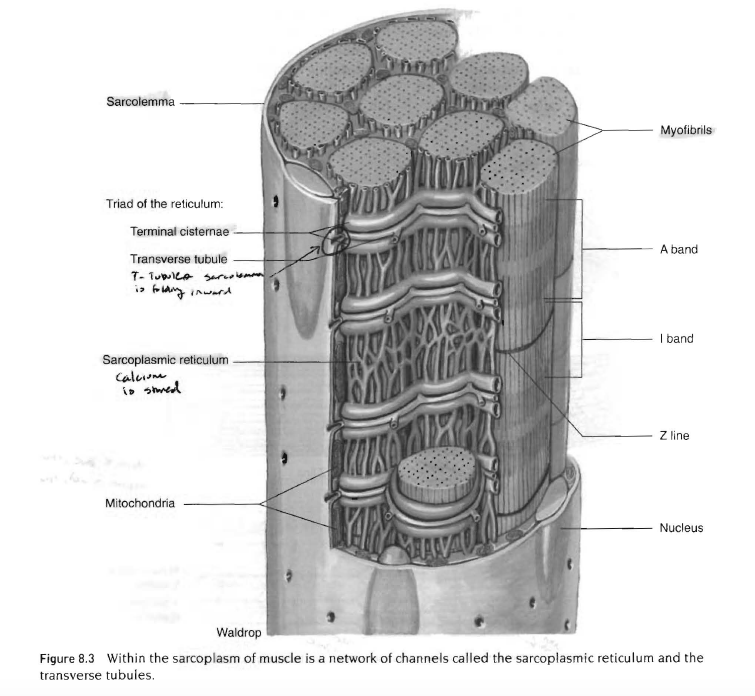
근섬유 - 500여개의 근원섬유의 연결
근초, 근형질세망, 위성세포
위성세포는 분화하지 않은 세포로서 근육의 성장과 보수에 중요한 역할을 함. 만약 근섬유가 부상 또는 질병에 의해 파괴되면, 세포분화로 모두 채워질 수 없음. 그렇지만 위성세포로부터 새로운 섬유가 형성되어 분화와 성장을 통하여 성숙된 근섬유를 형성하는 것은 가능함.
이와 더불어 위성세포는 근력훈련 시 기존의 근섬유에 핵을 제공하고 분화시킴으로써 근성장에 기여함. 근섬유에 핵의 숫자가 증가하면 단백질을 합성하는 능력을 증가시켜 근성장에 도움을 줌. 근섬유 성장을위한 추가적인 핵은 섬유가 개개의 핵에 세포부피의 일정한 비율을 유지하기 위해 사용하는 전략으로 핵주변 세포질 크기를 근핵(myonuclear)이라고 함. 근핵 영역이 생물학적으로 가지는 의미는 하나의 핵이 필요한 유전자 표현, 즉 단백질 생산을 오직 세포의 제한된 구역에서만 할 수 있다는 것임. 그러므로 지속적으로 근핵을 유지하기 위해서 골격근 섬유가 성장하는 동안 위성세포에서 얻은 새로운 핵이 합쳐지는 것임. 이어 안정된 근핵 영역을 유지하기 위해 핵은 위축되는 동안 섬유로부터 손실됨.
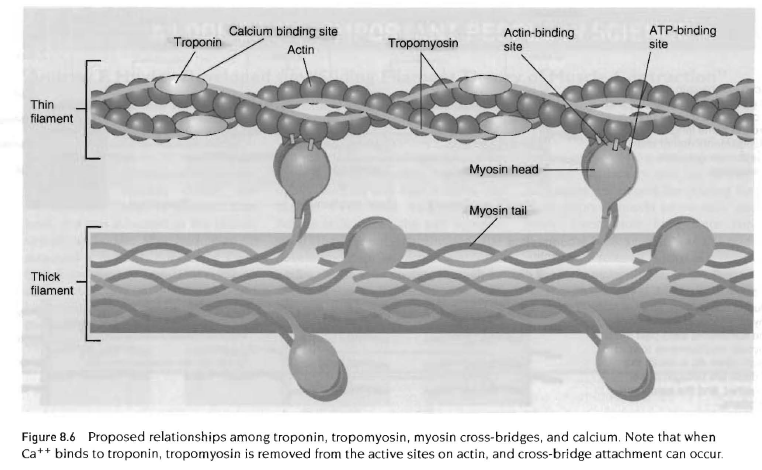
근초 아래에는 근형질(sarcoplasm)이 있는데 이것은 세포단백질, 기관, 근원섬유를 포함함. 근원섬유는 수축단백질을 포함하는 수많은 실같은 구조임. 근절은 마이오신과 액틴구조임. 액틴분자에는 또다른 단백질인 트로포닌과 트로포마이오신이 있음. 이 단백질은 근육의 작은 부분이지만 근수축 과정을 조절하는데 중요한 역할을 함.
근원섬유는 근절의 연결임.
근육의 근형질 내에는 각 근섬유를 둘러싸고 있는 그것과 평행하게 뻗어있는 막 통로 연결망이 있음. 이 통로를 근형질 세망(sarcoplasmic reciculum)이라하고, 칼슘 저장소로 근수축에 중요한 역할을 함. 또 다른 종류의 막 통로는 근섬유막내부로 연결되어 있는 가로세관(t-tubule)이라 부르며 근섬유의 근형질을 완전히 관통하고 있음. 이 가로세관은 종조(tenminal cisternae)라 불리는 두 근형질 세망사이를 관통하고 있음.
신경근 연접
각 골격근 세포는 신경세포로부터 오는 신경섬유가지에 연결됨. 이 신경세포는 운동신경이라 부르고 척수에서 밖으로 뻗어 있음. 운동신경과 그것이 지배하는 근섬유를 운동단위라고 함. 운동신경으로부터의 자극은 수축과정의 시초가 됨. 운동신경과 근세포가 만나는 곳을 신경근 연접이라고 함. 이 연접에서 호주머니 형태의 근섬유 막을 운동말판(motor end plate)라고 함.
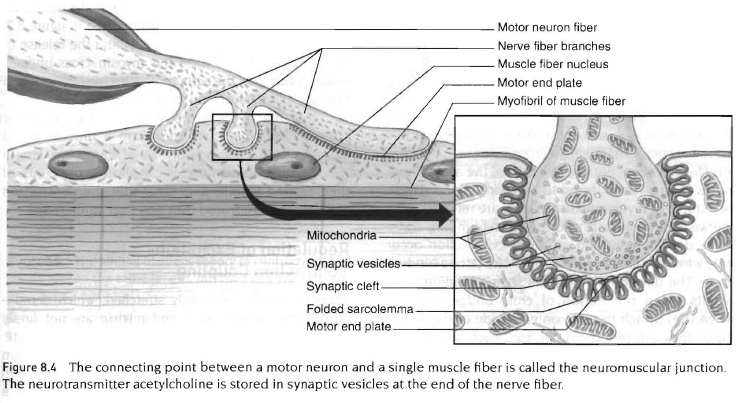
운동신경 말단은 근섬유와 물리학적인 접근을 하지 않으며 근신경 공간이라 불리는 연접틈(synaptic cleft)에 의해 분리됨. 신경자극이 운동신경의 말단에 다다를때 신경전달물질인 아세틸콜린이 방출되고 운동말판에 있는 수용체 부분과 결합하기 위해 공간틈세를 가로질러 확산됨. 이것은 나트륨에 대한 근섬유막의 투과력을 증가시키고, 말판전위(end-plate potential)라 부르는 탈분극의 결과를 가져옴. 말판전위는 항상 역치이상으로 충분히 커서 수축과정이 시작되는 신호가 됨.
근수축
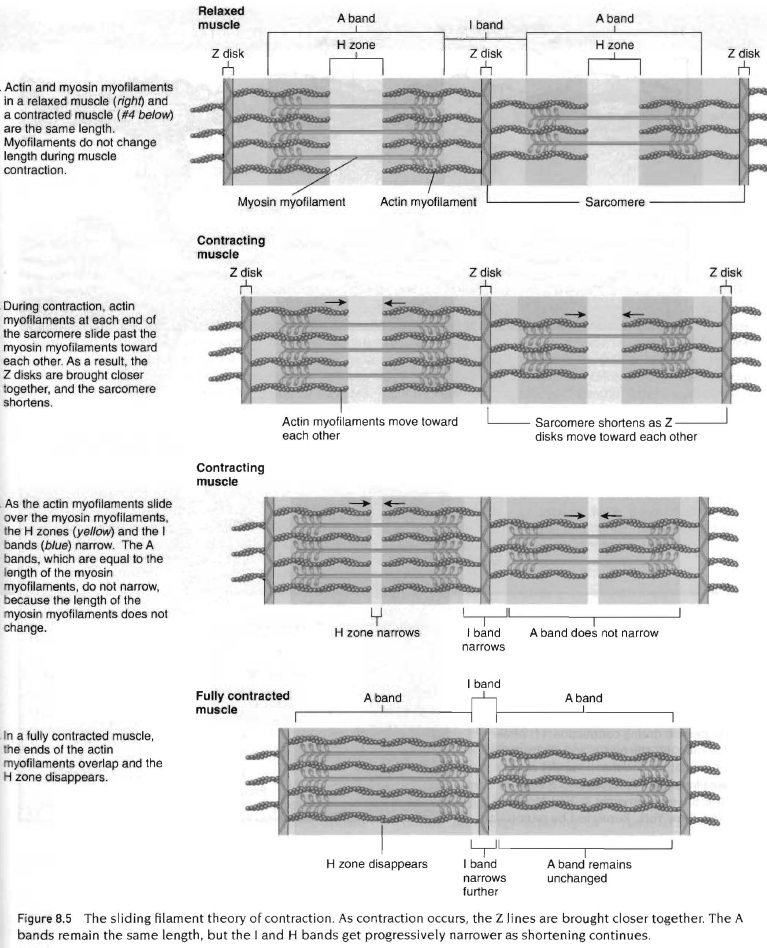
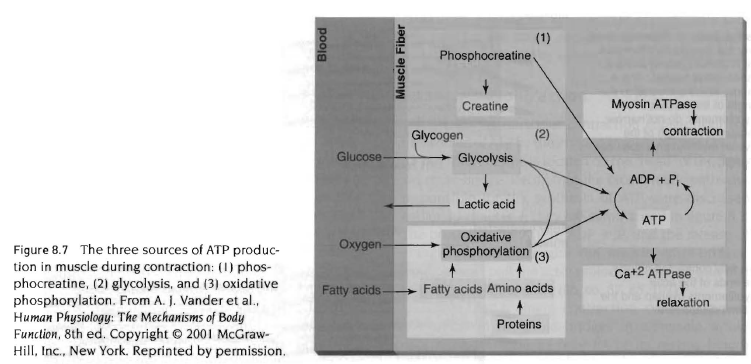
근수축은 많은 세포단백질과 에너지 생산체계에 관여하는 복잡한 과정임. 근수축의 최종결과로서 마이오신 위로 액틴이 미끄러져 근육을 짧게 만들고 근육에 장력이 발생함. sliding filament therory라고 함.
sliding filament therory
근수축의 단계적인 과정은 아래 그림참조. 액틴이 마이오신 위로 미끄러지으로써 근원섬유가 짧아져 근섬유가 수축되고 이는 Z판 사이의 거리가 줄어드는 결과를 초래함. 따라서 근육이 수축하는 과정을 이해하기 위해서는 근원섬유의 미시적인 이해가 필요함. 특히 마이오신 십자형 가교의 머리가 액틴 분자와 결합하도록 뻗어져 있다는 사실을 이해하는 것이 중요함.
액틴과 마이오신 근세사가 근육이 수축하는 동안 서로 가로질러 미끄러지는 것은 마이오신 머리가 액틴의 활동부위에 결합함으로써 가능하며 이때 수많은 마이오신 십자형 가교가 작용함. 근육이 수축하면서 힘이 발휘되면 십자형 가교는 강한 결속을 유지함. 강한 결속상태는 십자형 가교가 액틴에 부착되어 각 분절의 중앙으로 액틴을 잡아당김. 액틴이 마이오신 위로 잡아당겨지면서 근육의 길이가 짧아지고 장력이 발생함. 자극-수축결합(exitation-contraction copuling)이란 용어는 신경자극(활동전압)이 근육 막에 전달되어 십자형 가교에 의해 근육이 짧아지는 일련의 과정을 나타낸 것임. 이과정은 단계적으로 다음에 설명할 것이며 우선 근수축의 에너지원에 대해서 알아봄.
수축을 위한 에너지
근수축을 위한 에너지는 ATPase에 의한 ATP분해로 일어남. 이 효소는 마이오신 십자형 가교의 머리에 위치하고 있음. ATP재합성을 위한 생체에너지 체계는 3장에서 논의. ATP가 ADP +Pi로 분해되면서 방출된 에너지는 마이오신 십자형 가교를 활성화시키며 이것은 교대로 마이오신 위로 액틴분자를 끌어당겨서 근육을 짧게 함.
한번의 수축사이클 또는 근육내의 모든 십자형 가교의 끌어당김은 근육의 길이를 휴식시 길이의 1%정도만 짧게 함. 따라서 특정근육이 휴식시 길이의 60%까지 짧게 하기 위해서는 근수축 주기가 계속 반복되어야 함.
자극-수축결합 조절
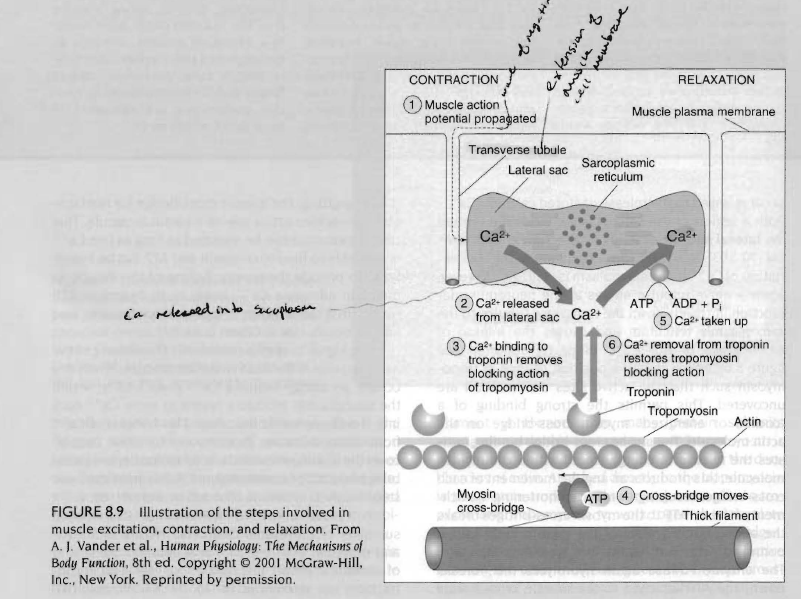
이완된 근육들은 쉽게 늘어져 있는데 이는 휴식시에 액틴과 마이오신이 확고히 부착되어 있지 않아서 서로 약하게 연결되어 있기 때문임. 액틴과 마이오신의 상호작용을 조절하면서 근수축을 조절하는 것은 무엇일까? 근육수축과정의 첫번째 단계는 신경근 연접에 이르는 신경자극으로 시작됨. 운동신경에서 나온 활동전압은 신경근 연접의 연접틈 공간에서 아세틸콜린을 방출함. 아세틸콜린은 근섬유막에 있는 수용체와 결합되고, 역치를 능가하는 말판전위를 가져와 근육세포의 탈분극을 이끔. 이 탈분극은 근섬유 속 깊에 있는 가로소관으로 전도됨. 활동전압이 근형질 세망에 도달되면 칼슘이 방출되고, 트로포닌에 결합하기 위해서 근육속으로 확산됨(칼슘-트로포닌 결합). 이 단계가 바로 근수축을 유발하는 단계임. 왜냐하면 근수축조절은 액틴과 마이오신의 상호작용을 관리함으로써 근수축을 조절하는 트로포닌과 트로포마이오신, 두 조절단백질의 기능때문임.
트로포닌과 트로포마이오신이 어떻게 수축을 조절하는지를 이해하기 위해 액틴, 트로포닌, 트로포마이오신의 해부학적 관계를 이해할 필요가 있음. 액틴세사는 2개의 열이 비틀어진 상태로 정렬되어 있고 작고 많은 단백질 하부구조로 형성됨. 트로포닌은 액틴의 2개 열사이 홈에 위치하고 있는 얇은 분자임. 트로포 마이오신에 직접적으로 부착되는 것은 트로포닌임. 이 정렬은 트로포닌과 트로포마이오신이 액틴과 마이오신 십자형 가교의 부착을 조절하는데 있어서 함께 작용함. 이완된 근육에서 트로포마이오신은 액틴분자의 활성부위를 차단하는데 근수축이 일어나기 위해서는 마이오신 십자형 가교가 부착되어야 함.(액틴-마이오신 헤드 결합) 근수축이 일어나도록 하기 위한 기저는 종조(terminal cisternae)라 불리는 측낭(lateral sac)에 저장된 칼슘이온을 방출하는 것과 관련이 있음. 휴식하고 있는 이완된 근육에서 근형질 내의 칼슘이온 농도는 매우 낮음.
그러나 신경자극이 신경근연접에 다다랐을대 이것은 근형질세망에서 가로소관으로 이동하여 칼슘방출을일으키며, 트로포 마이오신의 위치를 변화시켜 액틴의 활동부위가 노출되어 에너지를 갖고 있는 마이오신의 십자형 가교가 액틴에 강하게 결합할 수 있도록 함. 강한 십자형 가교는 마이오신 분자내에서 저장된 에너지를 방출시키는데 이는 십자형 가교의 운동을 일으켜 결국 근육을 짧게 함. 마이오신의 십자형 가교에 새로운 ATP부착은 액틴과 마이오신의 십자형 가교의 결합력을 약화시킴. 효소 ATPase는 다시 마이오신 십자형 가교에 부착되어 있는 ATP를 분해하여 액틴분자의 결합부위에 재부착을 위해서 마이오신 십자형 가교에 필요한 에너지를 공급함. 이러한 수축현상은 칼슘이 트로포닌에 자유롭게 결합하고 ATP가 산화되어 필요한 에너지를 공급하는 한 계속 반복됨. 부적절한 칼슘 수준 또는 높은 수소이온 때문에 근육이 항상성을 유지하지 못하면 십자형 가교의 움직임을 방해하여 피로가 발생하게 됨.
수축을 멈추는 신호는 신경근 연접에서 신경자극이 없어지는 것임. 이것이 일어날때 근형질 세망내에서 칼슘 펌프가 근형질 세망속으로 칼슘을 되돌리기 위해서는 에너지가 필요함. 트로포닌으로부터 칼슘의 이동은 트로포마이오신이 액틴분자의 결합부위를 덮어서 십자형 가교의 활동을 멈추게 함. 아래 그림은 근수축과 이온과 관련된 기본과정을 설명함.
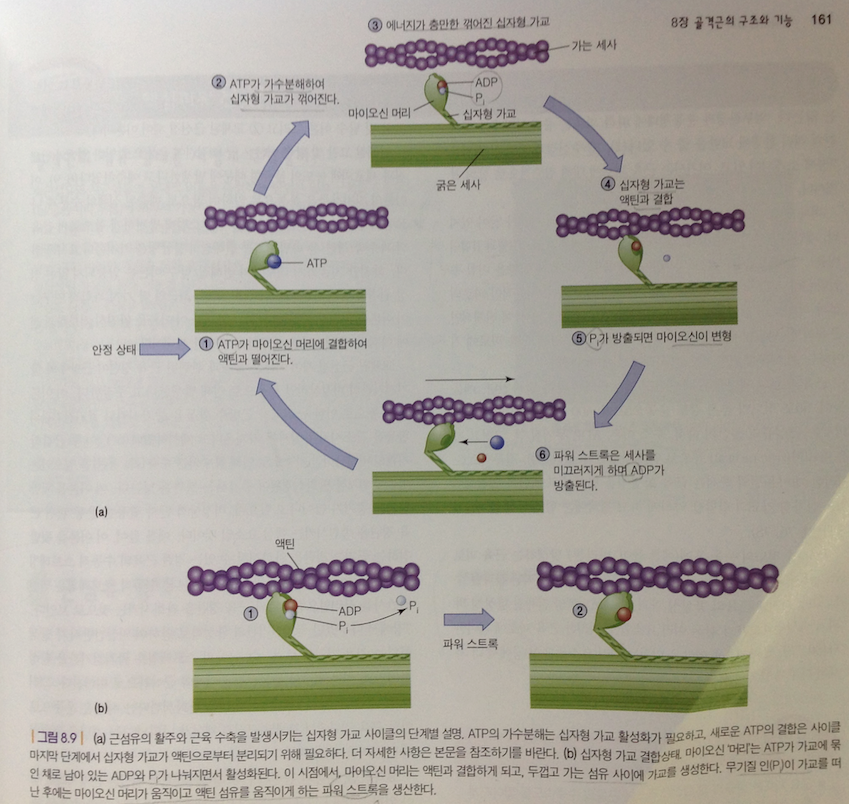
근수축 요약
1. 자극 단계
1) 운동신경의 활동전압은 신경근연접의 공간으로 아세틸콜린을 방출
2) 아세틸콜린은 운동말판의 수용체와 결합하여 탈분극을 유도하는 말판전위(EPP)를 야기시키며, 이는 근육내 가로소관으로 전도됨. 이렇게 탈분극이 일어나면 근형질세망으로부터 칼슘이온이 방출됨
2. 수축단계
1) 휴식단계에서는 마이오신 십자형가교는 액틴과 결합되지 않은 상태로 있음(힘생성 안함. 아래 그림 1단계 안정상태)
2) 탈분극이 근소포체에 도달하면 근형질세망으로부터 칼슘이온이 근형질로 방출됨. 칼슘이온은 트로포닌과 결합하고 트로포마이오신의 위치를 변형시켜 액틴의 활동부위를 노출시킴(그림 2단계). ATP분해는 활성화된 마이오신 십자형 가교를 형성하고(그림 3단계), 액틴의 활동부위와 강하게 결합함(그림 4단계)
3) 무기질 인은 마이오신 십자형 가교로부터 방출되고 활성화된 십자형 가교는 액틴을 잡아당김(그림 5단계)
4) 십자형 가교의 운동은 마이오신 십자형 가교에서 ATP가 ADP로 분해되는 에너지에 의해서 이루어짐.이 수축주기 시점에서 마이오신 십자형 가교는 액틴과 강한 결속을 이룸(그림 6단계).
5) 마이오신 십자형 가교의 ATP부착은 마이오신 십자형 가교의 강한 결속력을 약화시킴. 약화된 결속력은 ATP를 분해시켜 (ADP +Pi +에너지) 에너지를 발생시키고 이 에너지는 십자형 가교를 다시 활성시킴. (그림 1단계).
운동과 근육피로
고강도 또는 장시간 최대하 운동은 힘을 생산하는 근육의 능력을 저하시킴. 이러한 근육의 힘 산출량의 저하를 근 피로라고 함. 피로는 근육의 힘 생산량 감소와 근육의 수축속도를 짧아지게 함으로써 발생하는 근육힘의 힘 출력량 감소라고 정의됨.
운동으로 인한 근육피로에 대한 정확한 원인은 아직 논란의 여지가 남아 있지만, 개인의 신체적 건강상태, 영양균형상태, 근육의 조성(1형과 2형의 비율), 그리고 운동의 강도와 지속시간 등이 근피로에 영향을 미칠 수 있음. 더 나아가 근육피로의 원인에 상관없이 지속적인 피로의 결과 마이오신 십자형 가교에서의 낮은 힘생산으로 인하여 전체적인 근육의 힘 생산이 감소된다는 것임.
근육피로는 근섬유의 젖산축적, 수소이온, ADP, 무기질 인, 그리고 자유라디칼 등으로 발생할 수 있음. 이러한 대사물질의 축적은 근육 항상성을 방해하고, 액틴과 연결되어 있는 십자형 가교의 결합을 약하게하여 힘 생산이 줄어들게 됨. 자세한 것은 19장 참조.
운동과 관련된 근육경련
운동후 근육경련은 통증을 일으키며, 경련성의 불수의근 수축이 원인임. 1) 전해질 고갈 및 탈수이론 2)교체된 근신경 제어이론 두가지 이론이 존재함.
전해질 고갈 및 탈수이론은 근육경련이 운동으로 인한 혈액의 전해질과 세포외액 농도의 불균형때문에 발생한다고 생각함. 땀으로 배출된 수분과 나트륨의 손실은 운동신경 말단을 방전함으로써 근육의 과수축(경련)을 일으키는 전해질 불균형을 가져온다고 주장함.
교체된 근신경 제어이론은 운동과 관련된 근육경련이 근방추와 골지건기관의 비정상적인 활동으로 인해 발생한다고 주장함. 이 이론은 운동으로 인한 근피로가 근방추 활동을 증가시키고, 골지건기관의 활동을감소시키는 결과를 가져옴. 이 두 감각기관의 비정상적인 기능으로 인해 불수의근 수축(즉 경련)을 일으키는 척수에 위치한 발화신경원이 증가하는 결과를 낳음. 이 이론을 뒷받침하는 증거가 늘어나고 있으며, 비정상적인 반사활동은 운동관련 근육경련을 촉진시키는 핵심요인임.
즉 수동적 스트레칭은 역신전 반사를 일으켜 근육경련을 완화시키는 것으로 보임. 7장에서 다루었던 골지건기관의 활성화로 인하여 역신전 반사가 발생한다는 것을 기억. 즉 근육의 스트레칭은 골지건기관을 활성화시키고 이는 척수의 운동신경을 억제와 근이완으로 이어짐. 그러므로 경련근육의 스트레칭이 경련을 완화시킨다는 사실은 운동으로 인한 근육경련에서 교체된 근신경 제어가 나타난다는 개념을 뒷받침하는 근거를 제공함.
근섬유 형태
인간의 골격근은 각각 근섬유의 조직화학적 특성에 따라 여러종류로 구분할 수 있으며, 근섬유를 어떻게분류하려는가는 아래표를 참고. 특히 근섬유는 두가지 일반적인 범주로 분류됨.
1) 지근섬유 slow twitch fiber 2) 속근섬유 fast twitch fiber
지근섬유는 1형 단 한종류만이 인간의 근육에 존재하는 반면 속근섬유는 2형의 두가지 하위범주 2a,2x형이 존재함. 비록 몇면 근육군에서 속근섬유가 많거나 지근섬유의 비율이 높다고 알려져 있지만 대부분의 근육들은 두가지 형태의 비율이 균등하게 이루어져 있음. 개인의 골격근 섬유의 비율은 유전적 요인과 혈액속의 호르몬 수준, 운동형태에 따라 영향을 받음. 따라서 골격근을 구성하는 섬유비율은 파워와 지구력 경기에서 운동능력을 좌우하는 중요한 역할을 함.
골격근 섬유형태를 어떻게 구별할까? 속근섬유와 지근섬유의 상대적 비율을 측정하는 방법은 일부 근육을 추출하는 방법(biopsy)에 의해서 가능하며 근섬유의 조직화학적, 생화학적 분석에 의해서도 가능함. 일반적인 방법은 근섬유내의 마이오신 ATPase효소의 특별한 동위형태를 발견하는데 근거하여 근섬유를 세영역으로 구분하는 과정인 조직화학적 방법임. 이러한 기술을 통해 속근 섬유는 높은 농도의 ATPase를 함유하고 있어 화학적인 착색을 통해 어둡게 나타나며, 지근 섬유는 밝게 나타남. 속근섬유중에서 근섬유 2x형은 근섬유 2a형과 1형의 중간색깔임. 따라서 이 기술은 동시에 세 유형의 근섬유를 분류할 수 있음. 그림은 골격근막 단백질을 이용하여 조직화학적 염색을 한 후 근섬유 1형, 2a형, 그리고 2x형으로 볼 수 있음. 인간의 섬유형태에 대한 탐구에서 근생검법은 보통 한 근육군에서만 근육을 추출한다는 것임. 그러므로 한 근육군에서 추출된 샘플은 전체근육을 대표할 수 없는 것임. 더 복잡한 것은 섬유형태가 근육내 층을 이루고 있다는 것임. |
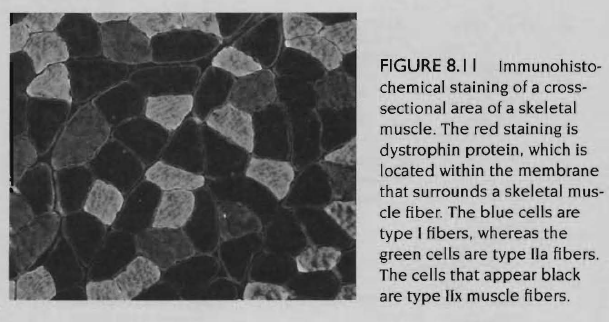
골격근의 생화학적 특성
근육의 세가지 주된 생화학적 특성은 근육기능에 중요한 산화능력(oxidative capacity), ATPase 효소의 동위형태, 근섬유내 풍부한 수축성 단백질임.
근섬유의 산화능력은 미토콘드리아의 수와 근섬유를 둘러싸고 있는 모세혈관의 수, 섬유내의 미오글로빈양에 의해 결정됨. 따라서 미토콘드리아 수가 많으면 유산소적인 ATP 에너지원을 많이 제공할 수 있는 능력이 뛰어남. 또한 근섬유를 에워싸고 있는 모세혈관이 많으면 근수축 시에 많은 양의 산소를 근섬유에 공급함. 미오글로빈은 혈액속의 헤모글로빈과 비슷하여 산소와 결합하며 세포와 미토콘드리아 사이에산소를 공급하는 역할을 함. 높은 함량의 미오글로빈은 모세혈관의 산소를 미토콘드리아에 많이 전달함으로써 근수축이 일어나는 근육에 많은 양의 에너지를 사용할 수 있게함. 결론적으로 근섬유가 많은 미토콘드리아, 미오글로빈, 모세혈관을 가지면 높은 유산소성 능력을 보유하게 되어 장기간 최대하 운동을해도 쉽게 피로하지 않음.

두번째로 중요한 근섬유의 생화학적 특성은 "마이오신 동형단백질 ATPase 효소의 활동성"임. 다양한 형태의 ATPase 효소의 동위형태가 존재하며 각각의 형태는 ATP를 활성화시키는 속도가 다름. 따라서 활동성 높은 ATPase 효소를 갖고 있는 근섬유는 ATP생성을 빠르게 촉진하여 근수축력을 강화시킬 수 있으며 낮은 ATPase는 근수축의 속도를 느리게 할 수 있음.
마지막으로 수축특성에 영향을 미치는 근섬유의 세번째 중요한 생화학적 특성은 근섬유에 풍부한 수축성 단백질(즉 액틴과 마이오신)임. 실재로 많은 양의 액틴과 마이오신을 함유한 섬유는 이러한 주요 수축성 단백질을 함유한 수치가 낮은 섬유보다 더 많은 힘을 생성함.
골격근의 수축특성
골격근의 수축특성은 네가지 운동형태가 중요함.
1) 최대근력발휘
2) 수축속도
3) 최대파워산출
4) 근섬유의 효율성
첫번째, 근섬유의 최대근력 발휘는 근횡단면적당 발휘하는 힘의 양을 나타냄.
둘째, 근섬유의 수축속도는 각 섬유의 최대수축속도를 측정하여 평가하며 최대수축속도는 근섬유가 최대속도로 수축할때를 나타냄. 근섬유는 십자형 가교의 움직임에 의해서 수축하기 때문에 최대 근수축 속도는 십자형 가교 주기의 비율에 의해 결정됨. 따라서 근섬유의 최대속도를 결정하는 생화학적 요인은 마이오신 ATPase 효소의 활동성임. 속근섬유처럼 높은 ATPase 효소를 갖고 있으면 최대수축속도 능력이 높은 반면, 지근섬유처럼 낮은 마이오신 ATPase를 갖고 있으면 수축속도 능력이 낮다.
세번째, 근섬유의 최대 파워산출. 근섬유의 파워는 수축생성과 수축속도에 의해 결정됨. 특히 근섬유의 최대파워산출은 힘생성과 수축속도를 곱한 값으로 정의됨. 따라서 높은 힘 생성능력은 가진 근섬유와 빠른 근수축속도는 높은 파워산출을 만든다.
넷째, 근섬유의 효율성. 근섬유의 효율성의 경제성을 평가하는데, 효율적인 근섬유는 일정한 운동을 하는데 필요한 에너지가 비효율적인 근섬유에 비해 적음.
근섬유 형태의 특성
1형 지근섬유, 2a, 2x형 속근섬유, 세가지 근섬유 유형이 인간의 골격근에 존재함.
지근섬유(type 1)
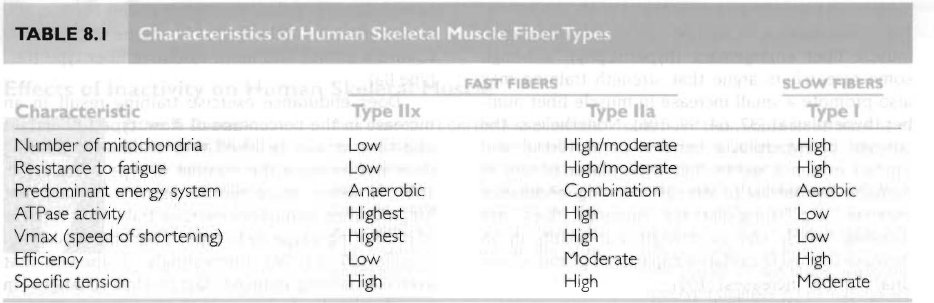
적색, 미오글로빈, 미토콘드리아, 산화효소, 모세혈관이 많음. 그래서 slow twich이고 피로에 높은 저항
최대수축속도는 느림. 낮은 마이오신 ATPase 활성.
지근섬유는 속근섬유보다 낮은 특이장력을 생산. 지근섬유가 속근섬유보다 단위면적당 적은 액틴과 미오신을 함유하고 있다는 것임. 근섬유에 의해 생성되는 힘의 양은 주어진 시간에 액틴과 마이오신의 상호교차결합의 수와 직접적으로 연관됨.
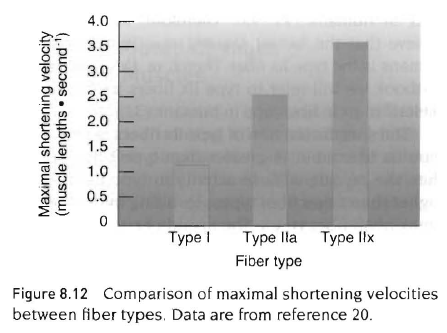
속근섬유(type 2a, 2x)
인간의 속근 섬유는 두가지 형태가 있음.
적은 미토콘드리아, 제한된 유산소성 대사능력, 쉽게 근피로.
속근섬유는 당원저장과 해당작용 효소가 풍부하여 무산소성 능력이 큼.
type 2x형, 2a형이 존재함.
운동을 하면 2x형이 2a형으로 전환함.

근섬유 형태와 수행능력
인간의 속근과 지근섬유 비율에 대한 몇가지 연구결과
1) 근섬유 분포에서 명백한 성별이나 나이차이는 없음.
2) 앉아서 일하는 직업을 가진 사람은 약 50%의 지근섬유를 가지고 있음
3) 파워운동 선수들은 속근섬유 비율이 높음. 지구성 운동선수들은 높은 지근섬유를 가지고 있음.
참고로 똑같이 성공적인 두명의 1만미터 달리기 선수가 갖고 있는 지근섬유 비율의 차이가 있을 수 있음. 예를들어 a선수는 70%의 지근섬유, b 선수는 85%의 지근섬유를 가지고 있음. 이런 관찰은 근섬유의 구성이 신체활동을 성공적으로 수행하기 위한 유일한 변인은 아니라는 것을 보여줌.
근육활동
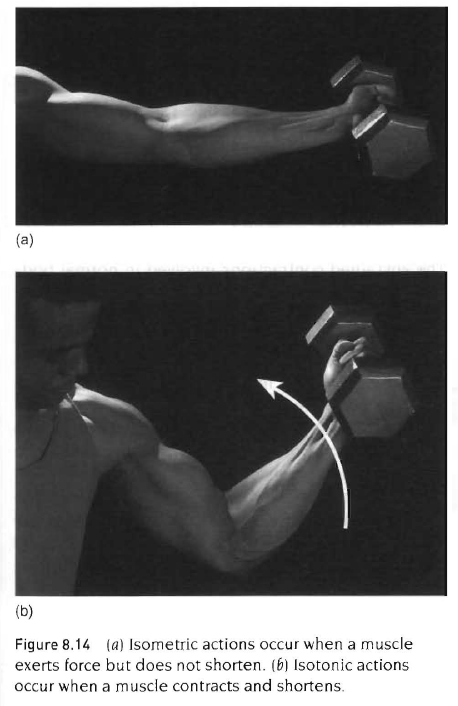
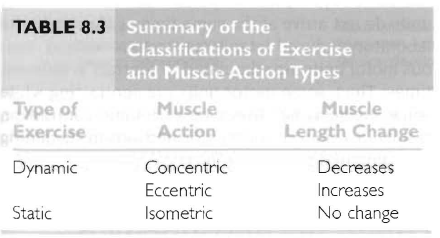
등척성 활동(isometric action)
등장성 활동(isotonic action) - 구심성 활동, 원심성 활동
근육에 스트레스 손상을 일으키는 활동은 원심성 근수축임.
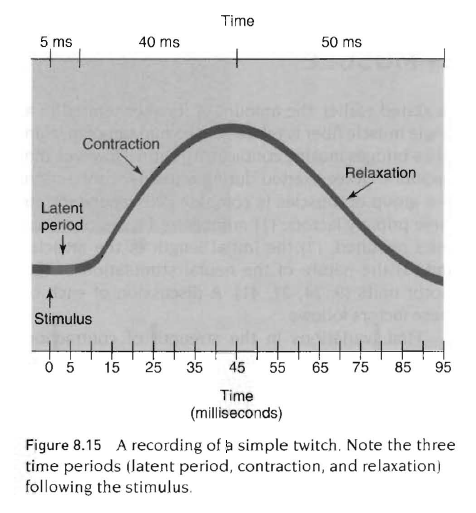
근수축과 이완속도
잠복, 수축, 이완으로 근수축을 구별할 수 있음.
여기서 delay in tension production, reaction time 개념과 lag in tension reduction개념이 나옴.
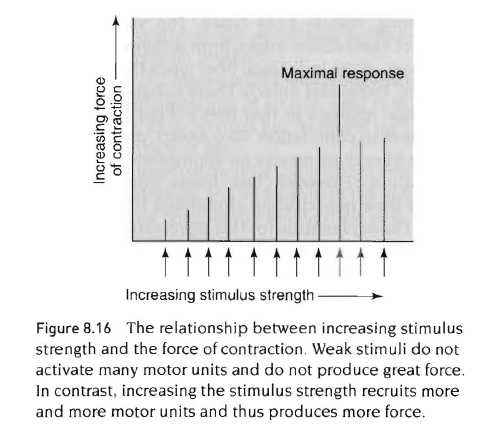
근육의 힘조절
하나의 근섬유내에서 발휘되는 힘의 양은 액틴과 접촉하는 마이오신 십자형 가교의 숫자와 관련이 있음.근육군 내에서 근육이 수축하는 동안 발휘되는 힘의 양은 복잡한 3개의 주요요인들에 의존함. 1) 동원된운동단위의 형태와 숫자 2) 근육의 초기 길이 3) 운동단위의 신경자극 특성임.
전체 근육내에서 수축력의 변화는 수축할때 자극을 받게 되는 근섬유의 숫자에 의존함. 오직 몇개의 운동단위가 동원된다면 그 힘은 작을 것임. 아래 그림은 이런 점을 보여주고 있음. 자극이 증가되면 수축하는 힘은 추가적인 운동단위의 동원때문에 증가함. 또한 속근섬유는 지근섬유보다 더 큰 장력을 발휘하므로 동원된 운동단위의 형태 또한 장력에 영향을 미침.
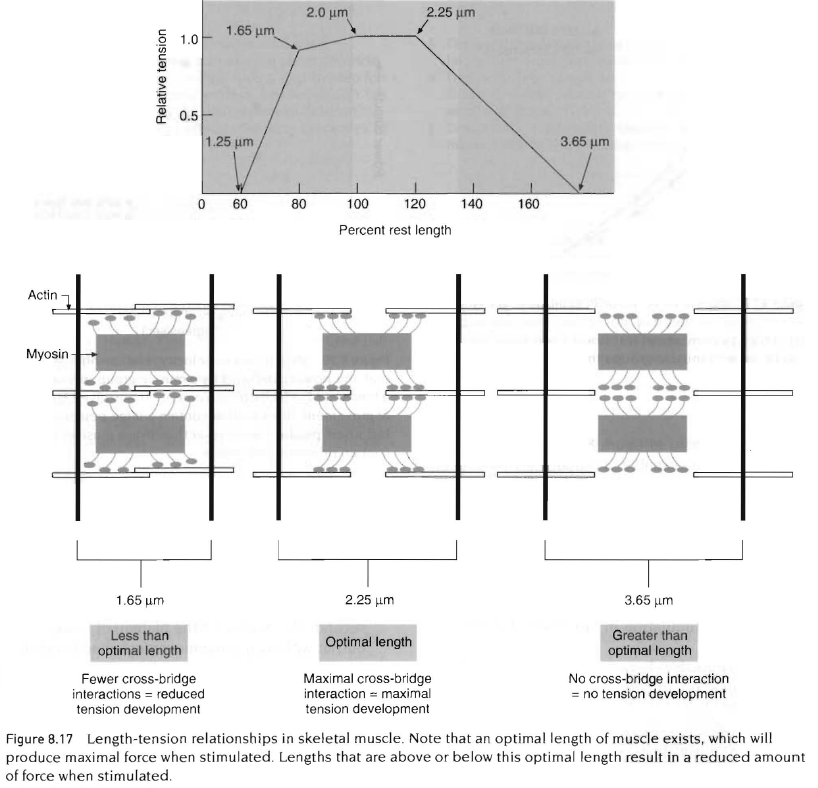
두번째 요인은 근육에서 발휘되는 힘은 수축시간과 근육의 초기 길이와 관련이 있음. 이것은 근섬유의 이상적 길이가 존재한다는 것임. 이상적인 길이의 존재에 대한 해석은 액틴과 마이오신이 서로 겹쳐지는것과 관련이 있음. 아래 그림 참조.
근육이 액틴과 마이오신의 겹쳐지는 부분이 없는 지점까지 늘어나게 되었을때, 십자형 가교는 부착될 수없기 때문에 장력을 더이상 발생할 수 없다는 점에 주목해야 함. 또 다른 극도의 경우는 근육이 휴식시간의 60%까지 짧아졌을때 Z판은 두꺼운 마이오신 세사에 매우 근접하게 되고 그 결과 제한된 추가적인 수축이 일어날 수 있음.
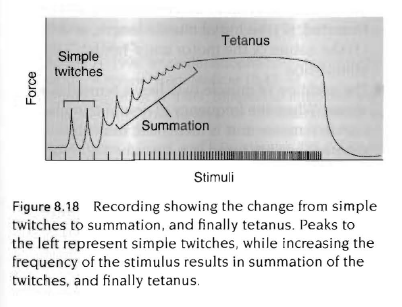
수축시 발휘되는 근력에 영향을 줄 수 있는 마지막 요인은 신경자극의 특성임. 실험적 상황에서 연구된 근육의 단축이 어떠한 근육기능인지 근본적인 특성에 몇가지 흥미로운 사실을 밝혀냄. 그러나 지속적인 수축을 포함하는 일반적인 신체의 움직임은 단축이 아님.
자극빈도가 증가하면 근육은 추가적으로 나타나는 자극과 힘 사이에서 이완하기 위한 시간을 가지지 않음. 이런 연속적인 수축 추가로 가중(summation)이 일어남. 더욱이 자극빈도가 증가하면 개개의 수축들은 중첩이 되어 하나의 강축이라고 불리는 수축을 유지함. 강축성 수축은 자극이 멈추거나 근육이 피로할때까지 지속될 것임.
근력생성에 영향을 주는 생리학적 요인에 대한 지금까지 논의에 덧붙여 노화와 질병은 힘을 생성할 수 있는 근육의 능력에 영향을 줄 수 있음. 실제로 노화가 근력생산을 감소시키는 근질량의 손실을 가져온다는 것은 잘 규명되어 있음.
힘-속도/파워-속도의 상관관계
대부분의 신체활동에서 근육의 힘은 운동범위를 통해서 적용됨. 이를테면 어떤 운동선수가 던지기를 할때 투척하는 물체를 놓기 전까지 운동범위에 따라서 힘이 적용됨. 얼마나 멀리 던질 수 있는가는 던지는 속도와 던지는 각도에 달려있음. 많은 운동경기에서 성공은 속도에 의존하기 때문에 근력과 움직임의 속도간 상관관계에 대한 기본적인 개념 몇가지를 인식하는 것이 중요함.
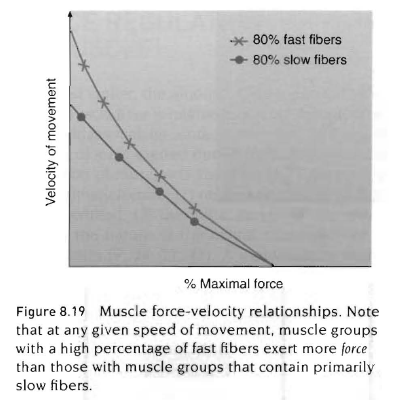
1) 근육이 절대적인 힘을 발휘할 때 근육의 속도 또는 움직임의 속도는 지근섬유가 많은 근육보다 속근섬유가 많은 근육에서 더 크다.
2) 근육수축 최대속도는 근육에 부과되는 저항이 가장 작은 시점임. 다시말하면 움직임의 최대속도는 가장 적은 부하에서 발생함.
힘-속도 곡선에 대한 생리학적 설명은 무엇인가? 힘의 생성은 액틴-마이오신 상호교차 수에 의존함. 액틴과 마이오신 상호교차연결에는 일정한 시간이 필요함. 그러므로 근육이 빠르게 수축할때(높은 속도의 운동), 액틴-마이오신 세사가 빠른 속도로 서로 이동함. 빠른 수축속도는 연결을 이룰 수 있는 상호 교차의 수를 감소하게 만들어 근력생산을 제한시킴. 반대로 근육수축속도가 감소함에 따라 더 많은 상호교차가 연결되고 근력생산이 증가함.
아래 그림은 속근섬유가 지근섬유보다 더 빠른 속도에서 큰 근력을 발휘한다는 것을 설명함. 생화학적 기전은 속근섬유가 지근섬유보다 더 높은 양의 활성화된 ATPase를 함유한다는 사실과 관련이 있음. 그러므로 ATP는 지근섬유와 비교할때 속근섬유에서 더 빠른 비율로 분해됨. 게다가 근형질세망에서 칼슘이 방출되는 것은 지근섬유보다 속근섬유의 신경자극 후에 더 빠름.
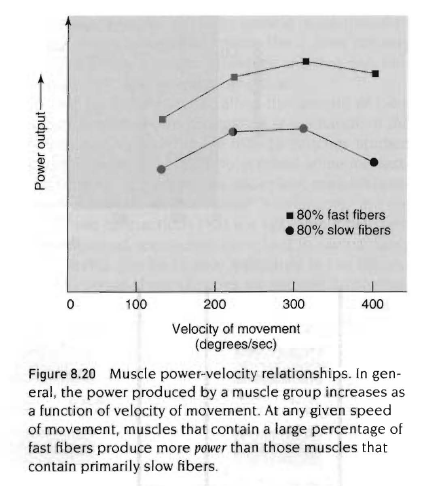
근섬유내 섬유-형태 분포는 파워-속도 곡선(위그림)에 영향을 줌. 근육을 통해서 발휘될 수 있는 최대파워는 높은 비율의 속근섬유를 함유한 근육에서 높은 비율의 지근섬유를 함유한 근육보다 더 큼. 힘-속도 곡선과 더불어 파워-속도 곡선도 두가지 중요한 점이 있음.
1) 주어진 운동속도에서 발생하는 최대파워는 높은 비율의 지근섬유를 가진 섬유보다는 높은 비율의 속근섬유를 함유한 근육에서 더 큼. 이런 차이는 앞에서 말한 속근섬유와 지근섬유 사이의 생화학적 차이때문임. 즉 높은 비율의 속근섬유를 가진 선수들은 주로 지근 섬유를 가진 선수보다 더 큰 파워를 냄
2) 근육을 통해서 발휘되는 최대파워는 200-300도/sec의 속도까지 운동속도를 증가시킴과 동시에 증가함. 운동속도의 증가와 함께 파워산출량의 고원상태에 대한 원인은 근력이 운동속도의 증가와 함께 감소하기 때문임. 그래서 주어진 근육군에서 가장 큰 파워를 산출할수 있는 적절한 운동속도가 있음.