- nature
- nature communications
- articles
- article
A single cell atlas of frozen shoulder capsule identifies features associated with inflammatory fibrosis resolution
- Article
- Open access
- Published: 19 February 2024
A single cell atlas of frozen shoulder capsule identifies features associated with inflammatory fibrosis resolution
- Michael T. H. Ng,
- Rowie Borst,
- Hamez Gacaferi,
- Sarah Davidson,
- Jessica E. Ackerman,
- Peter A. Johnson,
- Caio C. Machado,
- Ian Reekie,
- Moustafa Attar,
- Dylan Windell,
- Mariola Kurowska-Stolarska,
- Lucy MacDonald,
- Stefano Alivernini,
- Micon Garvilles,
- Kathrin Jansen,
- Ananya Bhalla,
- Angela Lee,
- James Charlesworth,
- Rajat Chowdhury,
- Paul Klenerman,
- Kate Powell,
- Carl-Philip Hackstein,
- ICECAP Consortium,
- Dominic Furniss,
- …
- Stephanie G. Dakin
Show authors
Nature Communications volume 15, Article number: 1394 (2024) Cite this article
Abstract
Frozen shoulder is a spontaneously self-resolving chronic inflammatory fibrotic human disease, which distinguishes the condition from most fibrotic diseases that are progressive and irreversible. Using single-cell analysis, we identify pro-inflammatory MERTKlowCD48+ macrophages and MERTK + LYVE1 + MRC1+ macrophages enriched for negative regulators of inflammation which co-exist in frozen shoulder capsule tissues. Micro-cultures of patient-derived cells identify integrin-mediated cell-matrix interactions between MERTK+ macrophages and pro-resolving DKK3+ and POSTN+ fibroblasts, suggesting that matrix remodelling plays a role in frozen shoulder resolution. Cross-tissue analysis reveals a shared gene expression cassette between shoulder capsule MERTK+ macrophages and a respective population enriched in synovial tissues of rheumatoid arthritis patients in disease remission, supporting the concept that MERTK+ macrophages mediate resolution of inflammation and fibrosis. Single-cell transcriptomic profiling and spatial analysis of human foetal shoulder tissues identify MERTK + LYVE1 + MRC1+ macrophages and DKK3+ and POSTN+ fibroblast populations analogous to those in frozen shoulder, suggesting that the template to resolve fibrosis is established during shoulder development. Crosstalk between MerTK+ macrophages and pro-resolving DKK3+ and POSTN+ fibroblasts could facilitate resolution of frozen shoulder, providing a basis for potential therapeutic resolution of persistent fibrotic diseases.
초록
동결견은
자연적으로 자가 해결되는 만성 염증성 섬유화 인간 질환으로,
진행성이고 비가역적인 대부분의 섬유화 질환과 구별됩니다.
단일 세포 분석을 통해
우리는 동결견 캡슐 조직에 공존하는
염증 촉진성 MERTKlowCD48+ 대식세포와 염증의 부정적 조절 인자가 풍부한
MERTK+LYVE1+MRC1+ 대식세포를 확인했습니다.
single-cell analysis
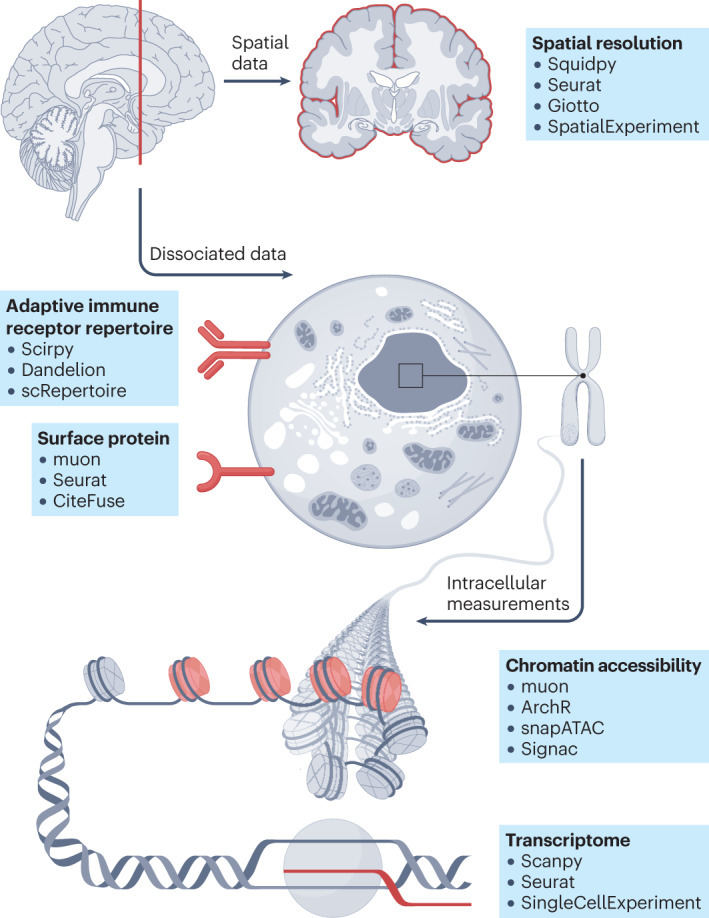
| 단일 세포 기술의 최근 발전으로 다양한 모달리티와 위치에 걸친 세포의 고처리량 분자 프로파일링이 가능해졌습니다. 단일 세포 전사체학 데이터는 이제 염색질 접근성, 표면 단백질 발현, 적응 면역 수용체 레퍼토리 프로파일링 및 공간 정보로 보완될 수 있습니다. 다양한 모달리티에 걸친 단일 세포 데이터의 가용성 증가는 분석가가 생물학적 통찰력을 도출하는 데 도움을 주는 새로운 계산 방법의 개발을 촉진했습니다. 이 분야가 성장함에 따라 방대한 도구와 분석 단계의 지형을 탐색하는 것이 점점 더 어려워지고 있습니다. 본 논문에서는 단일 및 다중 단일세포 분석 모달리티에 대한 독립적인 벤치마킹 연구를 종합하여 가장 일반적인 분석 단계에 대한 포괄적인 모범 사례 워크플로를 제안합니다. 독립적인 벤치마크가 없는 경우, 널리 사용되는 방법들을 검토하고 비교합니다. 본 논문은 단일세포 (다중)오믹스 분석 분야의 초보자에게 입문 지침이 되고, 고급 사용자에게는 최신 모범 사례를 안내합니다. |
| 단일 세포 RNA 시퀀싱(scRNA-seq) 기술은 전례 없는 규모와 해상도로 전사체 프로파일을 측정할 수 있게 함으로써 분자 생물학에 혁명을 일으켰습니다. 실험 기술의 발전은 대규모의 계산 방법 혁신을 촉진하여 현재 scRNA-seq 데이터를 분석하기 위해 1,400개 이상의 도구가 이용 가능하게 되었습니다1. Bioconductor2, Seurat3, Scanpy4와 같은 계산 프레임워크 및 소프트웨어 저장소는 방법 벤치마크와 모범 사례 워크플로우2,5,6과 함께 데이터 분석가가 이 영역을 탐색하고 분석 파이프라인을 구축할 수 있도록 지원합니다. 이러한 실험적 혁신과 계산적 혁신의 상호작용은 조직 세포 이질성을 밝혀내는 생물학적 획기적 발견을 가능하게 했습니다7,8. 그러나 scRNA-seq은 세포 기능과 신호전달을 지배하는 복잡한 조절 기전의 한 층만 포착합니다. 이를 보완하기 위해 염색질 접근성9, 표면 단백질10, T 세포 수용체(TCR)/B 세포 수용체(BCR) 레퍼토리11 및 공간적 위치12 등이 포함됩니다. 이를 통해 제2형 당뇨병 조절 시그니처13, 중증 급성 호흡기 증후군 코로나바이러스 2(SARS-CoV-2)에 대한 선천성14 및 적응성15 면역 체계의 조절 장애 반응, 그리고 공간적 해상도에서 종양 미세환경의 면역억제 효과에 대한 더 나은 이해16 등의 발견이 가능해졌습니다. 실험적 혁신으로 다양한 단일 세포 오믹스 모달리티를 위한 수많은 새로운 계산 도구가 개발되었으나, 모범 사례 워크플로우의 부재로 인해 방대한 신기술 도구 환경을 탐색하는 것이 어렵습니다. 또한, 이전에 scRNA-seq2,5,6,17에 대한 계산적 모범 사례 및 도구 권장 사항이 제시된 바 있으나, 이들은 구식이거나 불완전합니다. 본 논문에서는 단일 모달리티 및 다중 모달리티 단일 세포 데이터 분석의 다양한 단계를 안내하고 분석 시 주의사항과 권장사항을 논의한다(그림 1). 도구의 신생성이나 독립적 벤치마크 부재로 인해 모범 사례를 확정할 수 없는 경우, 널리 사용되는 도구와 커뮤니티 권장사항을 제시한다. 본 논문은 단일 워크플로 대신 모달리티별 섹션과 분석 단계 그룹으로 구성됩니다. 현대 단일 세포 분석에서는 작업의 다양성으로 인해 단일 워크플로가 거의 존재하지 않기 때문입니다. 추가 참고 자료로, 상세한 코드 예시, 분석 템플릿 및 계산 요구 사항 평가를 포함한 50개 이상의 장으로 구성된 보다 포괄적이고 정기적으로 업데이트되는(그러나 동료 검토를 거치지 않은) 단일 세포 최적 관행 온라인 책을 제공합니다. |





환자 유래 세포의 미세 배양을 통해
MERTK+ 대식세포와 염증 해결을 촉진하는 DKK3+ 및
POSTN+ 섬유아세포 사이의 인테그린 매개 세포-기질 상호작용을 확인하였으며,
이는 기질 재구성이 동결견 해결에 역할을 함을 시사한다.
교차 조직 분석을 통해
어깨 관절낭 MERTK+ 대식세포와 류마티스 관절염 환자의 관절막 조직에서 풍부하게 발견되는
특정 집단 사이에 공유되는 유전자 발현 카세트가 존재함을 확인하였으며,
이는 MERTK+ 대식세포가 염증 및 섬유화의 해결을 매개한다는 개념을 뒷받침합니다.
인간 태아 어깨 조직의 단일 세포 전사체 프로파일링 및 공간 분석을 통해
동결견에서 발견되는 것과 유사한 MERTK+ LYVE1+ MRC1+ 대식세포 및 DKK3+ 및 POSTN+ 섬유아세포 집단을 확인하여,
섬유증 해결을 위한 템플릿이 어깨 발달 과정에서 확립된다는 점을 시사한다.
MerTK+ 대식세포와 분해 촉진성 DKK3+ 및 POSTN+ 섬유아세포 간의 교신은
동결견의 해결을 촉진할 수 있으며,
이는 지속적인 섬유화 질환의 잠재적 치료적 해결을 위한 기초를 제공한다.
Similar content being viewed by others

CD200+ fibroblasts form a pro-resolving mesenchymal network in arthritis
Article 23 February 2024
Article Open access31 December 2022
Article 20 July 2023
Introduction
Intractable inflammatory fibrotic diseases affecting soft tissues including lung, liver, kidney and skin contribute to 45% of all-cause mortality1. Clinically effective therapies that promote resolution of fibrosis are currently lacking. Recent single-cell transcriptome sequencing (scRNA-seq) analyses identify multiple scar-associated cell lineages that populate fibrotic visceral tissues. For example, cirrhotic human livers show enrichment for pro-fibrotic pathways including TNFRSF12A, PDGFR and NOTCH signalling with the cirrhotic niche being comprised of TREM2 + CD9+ macrophages and ACKR1 + PLAVP+ endothelial cells2. In pulmonary fibrosis, a subset of alveolar macrophages from fibrotic lungs were found to express high levels of mediators such as IL1RN, MMP9 and SPP1 that are known drivers of inflammatory fibrosis3. Tissue-resident fibroblasts are likewise major constituents of the inflammatory fibrotic niche. Single-cell atlases of fibrotic human and murine lungs identify proliferating pathogenic myofibroblast subsets that are enriched for POSTN, CTHRC1, MFAP5 and ACTA23,4,5. Interactions between tissue-resident fibroblasts and pro-inflammatory macrophages induce fibroblast activation and irreversible deposition of fibrotic extracellular matrix1,6,7. These studies advance understanding of the biological processes culminating in solid organ fibrosis; however, the cellular basis by which fibrosis could resolve is not known.
서론
폐, 간, 신장 및 피부를 포함한 연조직에 영향을 미치는
난치성 염증성 섬유증 질환은 전체 사망 원인의 45%를 차지합니다1.
섬유증 해결을 촉진하는 임상적으로 효과적인 치료법은
현재 부족합니다.
최근 단일 세포 전사체 시퀀싱(scRNA-seq) 분석을 통해
섬유화된 내장 조직에 서식하는 여러 흉터 관련 세포 계통이 확인되었습니다.
예를 들어,
간경변 인간 간에서는 간경변성 미세환경을 구성하는
TREM2+ CD9+ 대식세포 및 ACKR1+ PLAVP+ 내피세포2와 함께
PDGFR 및 NOTCH 신호전달 경로가 풍부하게 관찰되며,
간경변 미세환경은 TREM2⁺CD9⁺ 대식세포와 ACKR1⁺PLAVP⁺ 내피세포로 구성된다2.
폐섬유증에서는
섬유화된 폐의 일부 폐포대식세포가
염증성 섬유증의 주요 유발인자로 알려진 IL1RN, MMP9 및 SPP1과 같은 매개체들을
고농도로 발현하는 것으로 확인되었다3.
조직 상주 섬유아세포 역시
염증성 섬유화 틈새의 주요 구성 요소이다.
인간 및 생쥐 섬유화 폐의 단일 세포 아틀라스는
POSTN, CTHRC1, MFAP5 및 ACTA2가 풍부한
증식성 병리학적 근섬유아세포 하위 집단을 확인하였다3,4,5.
조직 상주 섬유아세포와 염증성 대식세포 간의 상호작용은
섬유아세포 활성화를 유도하고
섬유성 세포외기질의 비가역적 침착을 유발한다1,6,7.
이러한 연구들은
고형 장기 섬유화로 이어지는 생물학적 과정에 대한 이해를 진전시켰으나,
섬유화가 해소될 수 있는 세포적 기전은 알려지지 않았다.
To understand how intransigent human fibrotic diseases might be therapeutically encouraged to resolve, we investigated frozen shoulder as an example of a localised chronic inflammatory fibrotic disease affecting the shoulder joint capsule that is normally self-limiting over time8. Frozen shoulder affects 10% of the working population, causing pain and disability through severely restricted shoulder joint motion9,10,11. Although characterised by localised fibrosis, a role for systemic factors in the pathogenesis of frozen shoulder cannot be excluded as it has been associated with both diabetes12 and Dupuytren’s disease13. Tissue biopsies from frozen shoulder patients with advanced-stage disease express inflammation-resolving proteins and show reduced expression of NFκB target genes compared to patients with persistent shoulder tendon tears14. These findings suggest that in frozen shoulder, the target tissues exhibit a resolving trajectory during advanced disease, which mirrors the typical clinical picture of reduced pain, stiffness and ultimate resolution over time. The unique biology of this under-investigated condition has the potential to unlock the cellular and molecular basis by which soft tissue inflammation resolves, persists or leads to fibrosis. Given that frozen shoulder is normally self-limiting, we hypothesised that the shoulder capsule is ‘primed to resolve’, and that the cellular interactions between major tissue-resident cell types including fibroblasts and macrophages (Mφ) might provide a resolving fibrotic niche conducive to restoring tissue homeostasis. Understanding this cellular basis for successful resolution could help provide the precise biological cues required to push persistent fibrotic diseases towards a resolving trajectory.
치료적으로 난치성 인간 섬유화 질환의 해결을 촉진하는 방법을 이해하기 위해,
우리는 일반적으로 시간이 지나면 자연적으로 제한되는8 어깨 관절낭에 영향을 미치는
국소적 만성 염증성 섬유화 질환의 예로서
동결견(frozen shoulder)을 연구하였다.
동결견은 근로 인구의 10%에게 영향을 미치며,
어깨 관절 운동의 심각한 제한을 통해 통증과 장애를 유발한다9,10,11.
국소성 섬유화로 특징지어지지만,
동결견의 병인에서 전신적 요인의 역할은 배제할 수 없는데,
이는 당뇨병12 및 뒤퓌트렌병13과 연관성이 확인되었기 때문이다.
진행된 단계의 동결견 환자 조직 생검에서는
지속적인 어깨 힘줄 파열 환자 대비 염증 해소 단백질 발현이 증가하고
NFκB 표적 유전자 발현이 감소하는 것으로 나타났다14.
이러한 결과는
동결견에서 표적 조직이 진행된 질환 단계에서 해결 경로를 보이며,
이는 시간이 지남에 따라 통증과 경직이 감소하고
궁극적으로 해결되는 전형적인 임상 양상을 반영함을 시사한다.
이처럼 연구가 부족한 질환의 독특한 생물학적 특성은
연부 조직 염증이 해결되거나 지속되거나 섬유화로 이어지는
세포적·분자적 기전을 규명할 잠재력을 지닌다.
동결견은
일반적으로 자가 제한적이라는 점을 고려할 때,
우리는 어깨 관절낭이 '해결을 위한 준비 상태'에 있으며,
섬유아세포와 대식세포(Mφ)를 포함한 주요 조직 상주 세포 유형 간의 세포 간 상호작용이
조직 항상성 회복에 유리한 해결성 섬유화 미세환경을 제공할 수 있다고 가정했습니다.
성공적인 해결의 이러한 세포적 기반을 이해하는 것은
지속성 섬유화 질환을 해결 경로로 이끌기 위해
필요한 정확한 생물학적 신호를 제공하는 데 도움이 될 수 있습니다.
As animal disease models do not accurately recapitulate frozen shoulder-affecting humans, we use scRNA-seq and multiparameter immunofluorescent histology to discover the cell types and microanatomical features that comprise the resolving fibrotic niche in frozen shoulder patient tissues. We identify a population of MERTK + Mφ residing in the capsule lining that are enriched for biological processes associated with the modulation of inflammation. Using micro-cultures of patient-derived cells, we demonstrate that MERTK + Mφ inhibit the inflammatory phenotype of capsular fibroblasts and reveal a cellular basis for resolution of frozen shoulder via integrin-mediated extracellular matrix remodelling. Cross-tissue analysis reveals that MERTK + Mφ in the shoulder capsule and in the knee synovium of rheumatoid arthritis (RA) patients share a common gene expression cassette. Finally, we demonstrate a possible embryonic origin for the cell types implicated in the resolution of frozen shoulder, suggesting that the cellular template for the resolution of fibrotic adult capsular disease is established during foetal development.
동물 질병 모델이 인간을 영향을 미치는 동결견을 정확히 재현하지 못하기 때문에,
우리는 scRNA-seq 및 다중 매개변수 면역형광 조직학을 사용하여
동결견 환자 조직에서 해결되는 섬유화 틈새를 구성하는 세포 유형과 미세해부학적 특징을 발견합니다.
우리는
캡슐 내막에 거주하며 염증 조절과 관련된 생물학적 과정이 풍부한
MERTK⁺ 대식세포 집단을 확인했습니다.
환자 유래 세포의 미세 배양을 통해,
MERTK⁺ 대식세포가
관절낭 섬유아세포의 염증성 표현형을 억제함을 입증하고,
인테그린 매개 세포외기질 재구성을 통한 동결견 해결의 세포적 기반을 밝힙니다.
교차 조직 분석 결과,
동결견 관절낭과 류마티스 관절염(RA) 환자의 슬관절 활막 내 MERTK⁺ 대식세포가
공통 유전자 발현 카세트를 공유함을 확인했습니다.
마지막으로,
동결견 해결에 관여하는 세포 유형의 배아 기원을 제시함으로써,
섬유화성 성인 관절낭 질환 해결의 세포적 틀이 태아 발달 기간에 형성됨을 시사합니다.
Results
Single-cell analysis of the resolving inflammatory fibrotic niche of the shoulder capsule
The shoulder capsule is comprised of a series of ligaments and a thin synovial layer encapsulating the glenohumeral joint. During frozen shoulder, the shoulder joint capsule becomes chronically inflamed and fibrotic, leading to significantly restricted range of motion. To understand the cellular basis by which inflammatory fibrosis associated with this condition ultimately resolves, we utilised well-phenotyped tissue biopsies from non-inflamed comparator and frozen shoulder patients to generate an atlas describing all the cell types comprising the adult shoulder capsule. Tissue biopsy samples were collected from the rotator interval of the shoulder capsule from male and female patients with a diagnosis of advanced stage frozen shoulder (duration of symptoms shown in Supplementary Data 1). Site and age-matched non-inflamed comparator tissue biopsies were collected from male and female patients undergoing shoulder arthroplasty. Histological examination of comparator capsule identified a clearly demarcated thin capsule lining and sparsely cellular underlying sub-lining region comprised of parallel-orientated collagen fibres (Fig. 1A). Conversely, tissue biopsies from frozen shoulder patients exhibited increased cellularity of both lining and sub-lining regions and increased vascularity (Fig. 1A). After integration with Harmony (version 0.1.0)15, Leiden clustering of 6818 cells isolated from 10 donor tissue biopsies (6 comparator, 4 frozen shoulder donors) revealed 5 major populations, each represented in cells from both comparator and diseased shoulder capsule donors (Fig. 1B and Figure S1A & B). Annotation of the stromal cells profiled from the adult shoulder capsule revealed that COL1A1 + COL3A1 + PDGFRB+ fibroblasts were the most abundant cell types followed by CD14 + CD68+ myeloid cells, CD3+T cells, PECAM1+ vascular endothelial cells and ACTA2+ mural cells (Fig. 1C & Figure S1C & D). Frozen shoulder patient tissues showed a trend towards increased proportions of lymphoid cells and PECAM+ vascular endothelial cells and a smaller increase in myeloid cells relative to comparator tissues. The proportion of fibroblasts was significantly reduced in frozen shoulder relative to comparator patient tissues (10% FDR; Fig. 1C). To support these observations at the protein level, we performed immunostaining to identify the major cell types comprising comparator and frozen shoulder patient tissues (Figure S1E). For quantification we segmented the resulting images into cell-associated ‘tiles’ by using the Voronoi algorithm to automatically place boundaries around the cell nuclei. On average, comparator shoulder capsule tissue samples comprised of 13.9% CD31+ vascular endothelial cells, 19.1% CD68+ macrophages and 67.0% DKK3+ fibroblasts. In comparison, frozen shoulder samples had increased relative proportions of CD31+ (22.6% vs 13.9%) and CD68+ (27.1% vs 19.1%) tiles, with a commensurate decrease in DKK3+ tile proportion (50.3% vs 67.0%). These proportional differences were not statistically significant (Figure S1E). In both conditions, CD31+ tiles were proportionally higher in the sub-lining region, whereas CD68+ tiles were proportionally higher in the lining region. Collectively, these data support our observations around the major cell types comprising comparator and frozen shoulder patient tissues, with the most abundant cell type being DKK3+ fibroblasts, followed by lining enriched CD68+ macrophages and sub-lining enriched CD31+ endothelial cells, in agreement with the scRNAseq data (Fig. 1B & C).
어깨 관절낭의 해결 중인 염증성 섬유화 미세환경에 대한 단일 세포 분석
어깨 관절낭은
일련의 인대와 견관절을 둘러싼 얇은 활막층으로 구성됩니다.
동결견 발생 시, 어깨 관절낭은
만성적으로 염증과 섬유화를 일으켜 운동 범위가 현저히 제한된다.
이 질환과 관련된 염증성 섬유화가
궁극적으로 해결되는 세포적 기전을 이해하기 위해,
우리는 비염증성 대조군 및 동결견 환자로부터 잘 표현형이 규명된 조직 생검을 활용하여
성인 어깨 관절낭을 구성하는 모든 세포 유형을 기술하는 아틀라스를 생성하였다.
조직 생검 샘플은 진행성 동결견 진단을 받은 남녀 환자(증상 지속 기간은 보충 자료 1 참조)의 어깨 관절낭 회전근개 간격에서 채취했습니다. 위치 및 연령이 일치하는 비염증성 대조군 조직 생검은 어깨 관절 치환술을 받는 남녀 환자에서 채취했습니다. 비교군 캡슐의 조직학적 검사를 통해 명확히 구분되는 얇은 캡슐 내막과 평행 배열된 콜라겐 섬유로 구성된 세포 밀도가 낮은 하부 내막 영역이 확인되었습니다(그림 1A). 반면, 동결견 환자의 조직 생검에서는 내막 및 하부 내막 영역 모두에서 세포 밀도 증가와 혈관 증가가 관찰되었습니다(그림 1A).
Harmony(버전 0.1.0)15와의 통합 후, 10명의 기증자 조직 생검(대조군 6명, 동결견 환자 4명)에서 분리된 6818개의 세포에 대한 라이덴 클러스터링 분석 결과, 대조군 및 병변 어깨 관절낭 기증자 모두의 세포에서 나타나는 5개의 주요 군집이 확인되었다(그림 1B 및 그림 S1A & B). 성인 어깨 관절낭에서 프로파일링된 기질 세포의 주석 분석 결과, COL1A1⁺ COL3A1⁺ PDGFRB⁺ 섬유아세포가 가장 풍부한 세포 유형이었으며, 그 다음으로 CD14⁺ CD68⁺ 골수계 세포, CD3⁺ T 세포, PECAM1⁺ 혈관 내피 세포 및 ACTA2⁺ 벽세포가 뒤를 이었다(그림 1C 및 그림 S1C & D).
동결견 환자 조직은
대조군 조직에 비해 림프구 세포와 PECAM+ 혈관 내피 세포의 비율이 증가하는 경향을 보였으며,
골수성 세포의 증가는 상대적으로 적었다.
섬유아세포의 비율은
대조군 환자 조직에 비해 동결견에서 유의하게 감소했다(10% FDR; 그림 1C).
이러한 관찰을 단백질 수준에서 뒷받침하기 위해,
대조군 및 동결견 환자 조직을 구성하는 주요 세포 유형을 확인하기 위한
면역염색을 수행했습니다(그림 S1E).
정량화를 위해, 결과 이미지를 세포 핵 주위에 자동으로 경계를 설정하는 보로노이 알고리즘을 사용하여 세포 관련 '타일'로 분할했습니다. 평균적으로 대조군 어깨 관절낭 조직 샘플은 CD31+ 혈관 내피 세포 13.9%, CD68+ 대식세포 19.1%, DKK3+ 섬유아세포 67.0%로 구성되었다. 이에 비해 동결견 샘플은 CD31+ (22.6% vs 13.9%) 및 CD68+ (27.1% vs 19.1%) 타일의 상대적 비율이 증가했으며, 이에 상응하여 DKK3+ 타일 비율은 감소했습니다 (50.3% vs 67.0%). 이러한 비율 차이는 통계적으로 유의하지 않았습니다(그림 S1E). 두 조건 모두에서 CD31+ 타일은 하부 내막 영역에서 비율이 더 높았고, CD68+ 타일은 내막 영역에서 비율이 더 높았습니다. 종합적으로, 이 데이터는 비교군 및 동결견 환자 조직을 구성하는 주요 세포 유형에 대한 우리의 관찰을 뒷받침하며, 가장 풍부한 세포 유형은 DKK3+ 섬유아세포이며, 그 다음으로 내막에 풍부한 CD68+ 대식세포와 하부 내막에 풍부한 CD31+ 내피세포가 이어집니다. 이는 단일세포 RNA 시퀀싱(scRNAseq) 데이터와 일치합니다(그림 1B 및 C).
Fig. 1: Cell types comprising the resolving inflammatory fibrotic niche.
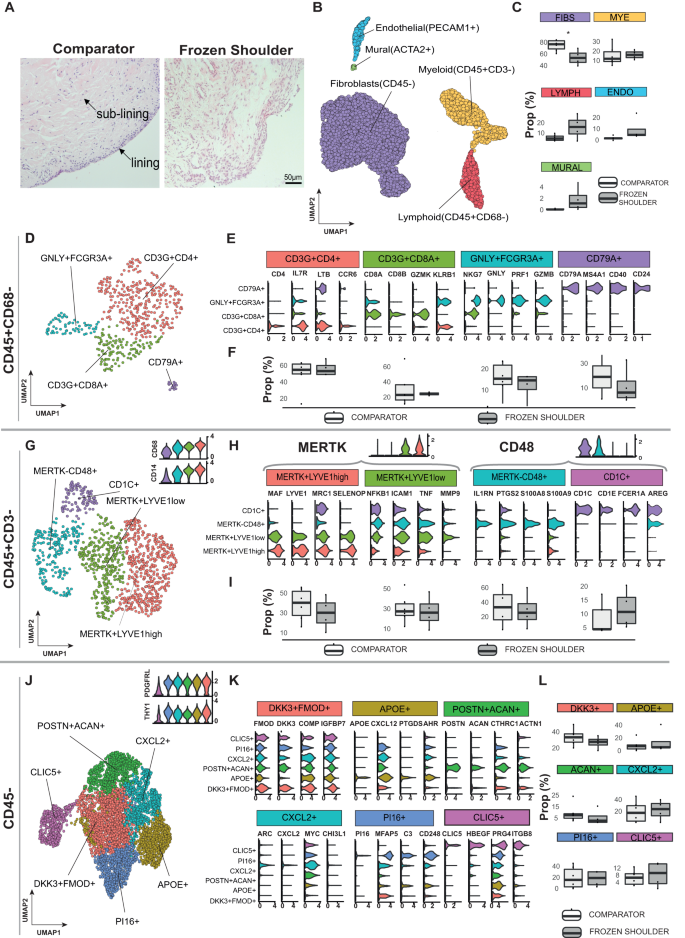
A Representative images showing Haematoxylin & Eosin staining of sections of comparator and frozen shoulder patient tissues. Distinct lining and sub-lining regions of the capsule are identified, frozen shoulder tissue sections show increased cellularity and vascularity relative to comparator tissues. Nuclear counterstain is violet, scale bar=50μm. scRNA-seq analysis of adult shoulder capsule from tissue biopsy samples collected from comparator (n = 6) and frozen shoulder (n = 4) patient donors. B UMAP shows the major cell types identified (resolution = 0.4) C The boxplots show the relative frequencies of the cell types in comparator and frozen shoulder patient tissues highlighting credible differences in the proportion of fibroblasts between these sample types (indicated by *). D The 4 identified lymphoid clusters (resolution = 0.2). E Violin plots of selected lymphoid cluster marker genes. F Relative frequencies of lymphoid clusters in comparator and frozen shoulder patient tissues. G The 4 identified myeloid clusters (resolution = 0.2). H Selected myeloid cluster marker genes. I Relative frequencies of myeloid clusters in comparator and frozen shoulder patient tissues. J The 6 identified fibroblast clusters (resolution = 0.3). K Selected fibroblast cluster marker genes. L The relative frequencies of the fibroblast clusters in comparator and frozen shoulder patient tissues. The credibility of differences in composition between the frozen shoulder and comparator samples was determined for each clusters C, F, I and L with scCODA74 (10% FDR). Violin plots in E, H, K show log-normalized expression values of selected cluster marker genes. Only significant cluster marker genes are shown (two-sided Wilcoxon tests, BH adjusted P values < 0.05). Box plots in C, F, I and L show median as centre, lower and upper quartiles as box limits, and whiskers with a length of 1.5 IQR.
Immune cell atlas of frozen and comparator human shoulder capsule
A separate analysis of the lymphoid cells revealed two distinct CD3+T cell clusters, NK cell and B cell clusters (Fig. 1D, Figure S2A & B, Supplementary Data 2). The CD4+ cluster expressed IL7R, CCR6, TIMP1, CD40LG and LTB; the CD8A/B+ cluster expressed GZMK and KLRB1 (Fig. 1E, Figure S2C). Within the lymphoid cell subset, the relative proportions of the T cell and NK cell clusters was similar between comparator and frozen shoulder patient tissues while the B cells showed a slight but non-credible decrease in frequency in the frozen shoulder samples (Fig. 1F). Geneset over-representation analysis using gene ontology (GO) Biological Processes (BP) in CD4+ cells revealed enrichment for ‘T helper cell differentiation’ and ‘regulation of leukocyte proliferation’; CD8+ cells were enriched for leukocyte and natural killer cell-mediated cytotoxic processes (Figure S2D). Small clusters of NKG7 + NK cells expressing cytotoxic molecules including PFR1, GNLY, GZMB and CD79A B cells were also identified (Fig. 1E & Fig. S2D).
A separate analysis of the myeloid cells identified four clusters in adult shoulder capsule tissues (Fig. 1G, Figure S3A & B, Supplementary Data 2). They included two MERTK + Mφ clusters; a LYVE1high cluster expressing MRC1, MAF, COLEC12 and a second LYVE1low cluster expressing FCGR3A, ICAM, TNF, MMP9 (Fig. 1H). A MERTKlowCD48high macrophage cluster expressing PTGS2, IL1RN and S100A8 and a small cluster of CD48+ monocyte-derived dendritic cells (CD1C, AREG, FCER1A) were also identified (Fig. 1H). The relative proportions of these myeloid clusters did not significantly differ between comparator and frozen shoulder patient tissues (Fig. 1I). To investigate possible changes in macrophage phenotype between frozen shoulder and comparator tissues, we performed pseudo-bulk based differential gene expression analyses Figure S3C–E (Supplementary Data 3). Grouping of all myeloid cells revealed significantly elevated SPP1 expression in comparator relative to frozen shoulder tissues (BH adjusted P = 2.32 × 10−9; DESeq2 analysis; Figure S3C, Supplementary Data 3). Within the macrophage clusters, we detected n = 59 genes in the MERTK+LYVE1lowMφ, n = 92 genes in the MERTKlowCD48highMφ and n = 8 genes in the MERTK+LYVE1highMφ as differentially expressed between the frozen shoulder and comparator patient tissues. Of note, MERTK+LYVE1lowMφ isolated from frozen shoulder patient tissues had significantly higher expression of CSF2RA, NFKB1, ITGAX and TNFRSF1B (BH adjusted p < 0.001; DESeq2 analysis; Figure S3D, Figure S4A, Supplementary Data 3). MERTK+LYVE1high Mφ isolated from frozen shoulder patient tissues showed little difference to those from comparators but had significantly lower expression of SPP1 (BH adjusted p < 0.001; DESeq2 analysis; Figure S4A, Supplementary Data 3). MERTKlowCD48high Mφ isolated from frozen shoulder patient tissues had significantly higher expression of CSF2RA, IL1A and IL1B (BH adjusted p < 0.05; DESeq2 analysis; Figure S4A, Supplementary Data 3). The expression of selected pro-inflammatory and immunomodulatory genes in the myeloid cell clusters is shown in Figure S4A & B. We noted that the MERTK+LYVE1highMφ cluster showed robust expression of immunomodulatory genes including CMKLR1, LYVE1, MRC1 and TGFB family genes in the frozen shoulder samples. Collectively, these findings demonstrate that comparator and frozen shoulder patient tissues are comprised of heterogeneous myeloid cell populations that have a modified transcriptional phenotype in frozen shoulder patients.
Stromal cell atlas of frozen and comparator human shoulder capsule
Fibroblasts exist as distinct subsets with diverse roles in tissue homeostasis, inflammation and fibrosis16,17,18,19. They constitute the major cell type of the shoulder joint capsule, yet their precise phenotypes have not been described in this tissue. We therefore performed a targeted analysis of the COL1A1 + COL3A1 + PDGFRB+ fibroblasts (Fig. 1J). After integration, six clusters were identified (Figure S5A & B). These included five THY1 + PDGFRL+ clusters: (i) “DKK3 + FMOD+” cells, (ii) “APOE+” cells expressing CXCL12, (iii) “POSTN + ACAN+” cells, (iv) “CXCL2+” cells expressing ARC, (v) “PI16+” sub-population expressing MFAP5. In addition, we found a THY1lowPDGFRLlow “CLIC5+” cluster that also expressed HBEGF (Fig. 1K, Figure S5C, Supplementary Data 2). The relative proportions of fibroblast clusters did not significantly differ between comparator and frozen shoulder patient tissues (Fig. 1L). Geneset over-representation analysis revealed significant over-representations for the gene ontology (GO) biological processes related to collagen fibril and extracellular matrix organisation and ‘response to mechanical stimulus’ (DKK3 + FMOD+ and POSTN + ACAN+ clusters), chemotaxis (APOE+ cluster), apoptotic processes (CXCL2+ cluster), complement activation, humoral immune response and angiogenesis (PI16+ cluster) and complement activation, TNF production and ion channel binding (CLIC5+ cluster) (Figure S5D). Pseudobulk-level differential expression analysis of fibroblasts in frozen shoulder relative to comparator patient tissues is shown in Figure S5E. Of note, CLIC5, CSF1, ITGA8, SYNOP2 and CXCL1 showed significantly higher expression in the fibroblasts from frozen shoulder patient tissues (BH adjusted p < 0.01; DESeq2 analysis; Figure S4C & D, Figure S5E and Supplementary Data 4). Taken together, these findings suggest that comparator and frozen shoulder patient tissues are comprised of distinct fibroblast sub-populations which are involved in shaping the extracellular environment and the regulation of local tissue inflammation.
Spatial topography of the resolving inflammatory fibrotic niche
We next used immunofluorescence confocal microscopy to confirm the presence and to characterise the microanatomical niches of cell populations identified by single-cell analysis. Multiplex immunostaining using an extended panel of myeloid markers localised MerTK+Mφ subsets including LYVE1high & LYVE1lowMφ to the capsule lining (Fig. 2A). A cassette of protein markers further validated these distinct clusters including LYVE1, CD163, MRC1 (LYVE1high cluster) and FOLR2, ICAM1, CD83, FCGR3A (LYVE1low cluster), located in the capsule lining region, this lining region was expanded in frozen shoulder patient tissues (Fig. 2A, Figure S3F & Fig. S3G). MerTKlowCD48 + Mφ expressing PTGS2, IL1RA and S100A8 were identified in both lining and sub-lining regions and were abundant in frozen shoulder patient tissues (Fig. 2A, Figure S3F). Co-expression of Ki67 and CD163 support the proliferative phenotype of macrophages in frozen shoulder patient tissues (Figure S3H).
Fig. 2: Spatial topography of the resolving fibrotic niche.
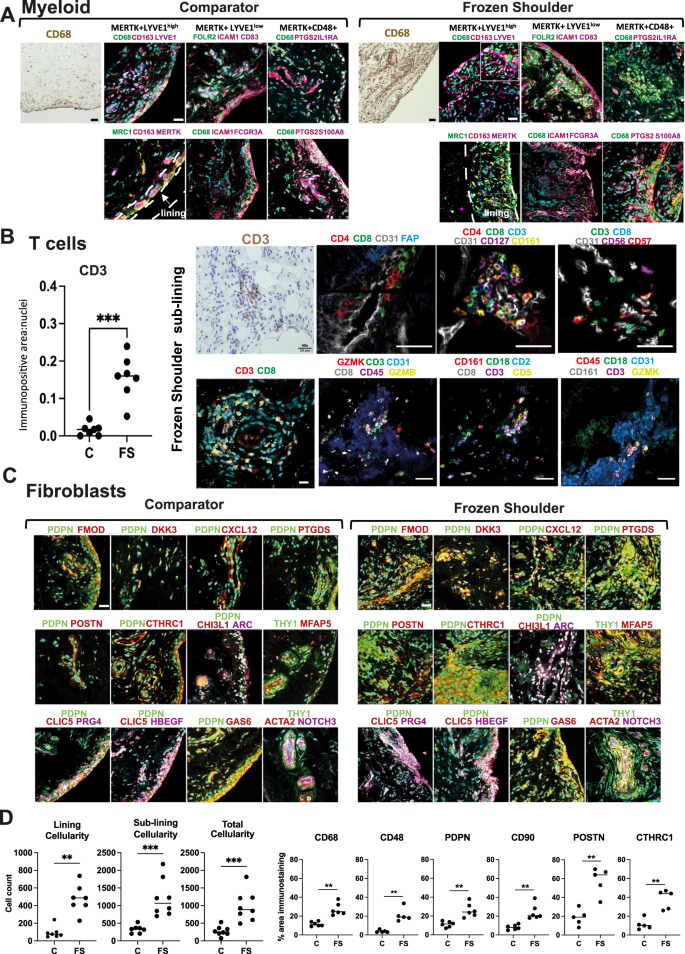
A Panel shows representative confocal images of immunostaining for CD68 and a wider panel of macrophage markers confirming the topographical niches of MerTK+LYVE1high, MerTK+LYVE1low and MerTKlowCD48+ macrophage subsets in sections of comparator and frozen shoulder patient tissues. MerTK + LYVE1highMRC1 + CD163+ and MerTK + LYVE1lowFOLR2 + ICAM1 + CD83 + FCGR3A+ subsets predominate in the capsule lining; triple positive cells are highlighted with arrow heads (Figure S3G). MerTKlowCD48 + PTGS2 + S100A8 + IL1RA+ macrophages occupy lining and sub-lining regions. Cyan represents POPO-1 nuclear counterstain, scale bar = 20 μm. B Graph shows quantitative analysis of immunostaining for CD3 in comparator (C, n = 7 donors) and frozen shoulder (FS n = 7 donors) patient tissue sections, pooled from 3 independent experiments, statistically significant differences were calculated using a two-sided Mann-Whitney test (P = 0.0006). Panel shows representative ChipCytometry images of T cells in sections of frozen shoulder patient tissues, residing adjacent to vascular endothelium (CD31+). Panels show staining combinations for CD4 + T cells (CD127+), CD8+ T cells (CD161 + GZMK+) and NK cells (CD56 + GZMB+), nuclei counterstained cyan/blue, scale bar = 50 μm. C Representative confocal images showing labelling for a cassette of fibroblast markers confirming the topographical niches of identified fibroblast sub-populations from Fig. 1J in sections of comparator and frozen shoulder patient tissues. Sub-population markers include DKK3 + FMOD+, CXCL12 + PTGDS+, POSTN + CTHRC1+, ARC+CHI3L1+, MFAP5+ and CLIC5 + HBEGF + PRG4+. Cyan represents POPO-1 nuclear counterstain, scale bar = 20 μm. D Graphs show quantitative analysis (QA) of total cellularity and cellularity localised to lining and sub-lining regions. Data generated using tissue sections derived from a minimum of 7 comparator and 7 frozen shoulder donors, pooled from 3 independent experiments. Statistically significant differences were calculated using a two-sided Mann-Whitney test for lining cellularity (P = 0.001), sub-lining cellularity (P = 0.0003), total cellularity (P = 0.0003). QA analysis of macrophage markers in sections of tissue biopsies collected from comparator (C) and frozen shoulder (FS) patient donors including CD68 (n = 6 comparator and n = 6 frozen shoulder donors, P = 0.002) & CD48 (n = 5 comparator and n = 5 frozen shoulder donors, P = 0.008). QA for markers of fibroblast activation PDPN (P = 0.002) & CD90 (P = 0.002), (n = 6 comparator and n = 6 frozen shoulder donors for each marker). QA for matrix-associated markers POSTN (P = 0.008) and CTHRC1 (P = 0.008), (n = 6 comparator and n = 6 frozen shoulder donors for each marker). All immunostaining data were pooled from 4 independent experiments. Statistically significant differences were calculated using two-sided Mann-Whitney tests. Bars represent median values. ***P < 0.001, **P < 0.01.
Quantitative analysis of immunopositive staining revealed an increased number of CD3+ cells in frozen shoulder relative to comparator patient tissues (Fig. 2B, P = 0.0006). CD3 + T cells resided adjacent to CD31+ vascular endothelium within the capsule sub-lining (Fig. 2B). Immunostaining confirmed the presence of CD4+, CD8+ and NK cells in frozen shoulder patient tissues, validating CD127 (CD4+ cluster), GZMB, GZMK, CD161 (CD8+ cluster) and CD56 + CD57+ (NK cell) populations (Fig. 2B).
Capsular fibroblasts also occupied distinct microanatomical niches. PDPN + PDGFRL+ fibroblasts populated lining and sub-lining regions, THY1+ (CD90 + ) fibroblasts predominated in the sub-lining adjacent to vascular endothelium (Figure S5F). CXCL12+, POSTN+ and MFAP5+ (PI16+ cluster) fibroblasts localised to sub-lining regions adjacent to blood vessels, DKK3 + FMOD+ and ARC+ (CXCL2+ cluster) fibroblast subsets were identified in both lining and sub-lining regions, CLIC5 + HBEGF+ fibroblasts mapped exclusively to the capsule lining (Fig. 2C). Mural cells (THY1 + ACTA2 + NOTCH3+) resided adjacent to vascular endothelium (Fig. 2C). Isotype control staining of capsular tissues is shown in Figure S6A–D. A summary of the cell types and microanatomy of the resolving inflammatory fibrotic niche is shown in Figure S6E.
Quantitative analysis of histological tissue sections identified increased cellularity across lining, sub-lining and combined regions of frozen shoulder patient tissues (P = 0.001, P = 0.0003 and P = 0.0003 respectively, Fig. 2D). Immunopositive staining for markers of tissue resident macrophages (CD68, P = 0.002), pro-inflammatory macrophages (CD48, P = 0.008), fibroblast activation markers (PDPN and CD90, P = 0.002 respectively) and matrix associated proteins (POSTN, CTHRC1, P = 0.008 respectively) were also increased in frozen shoulder relative to comparator patient tissues (Fig. 2D).
Capsular MERTKhigh macrophages have a modulatory phenotype
To identify candidate myeloid populations implicated in restoring homeostasis during advanced-stage frozen shoulder, we further investigated the phenotype and biological processes active in these cells. MERTK+LYVE1high macrophages expressed MAF, COLEC12 and SELENOP (Fig. 3A). The marker genes for this cluster were significantly over-represented for biological processes concerned with the regulation of humoral immune responses, regulation of complement activation, acute inflammatory response, and receptor-mediated endocytosis (Fig. 3B). Both MERTK+LYVE1high and MERTK+LYVE1low macrophage clusters expressed CD163, FOLR2 and MRC1 (Fig. 3C). The MERTK+LYVE1low cluster exhibited a mixed phenotype also expressing ICAM1, MMP9 and TNF (Fig. 3A & C). Biological processes significantly over-represented in the marker genes for this cluster included regulation of wound healing, cellular response to glucocorticoids and leucocyte chemotaxis (Fig. 3B). Significant over-representations in MERTKlowCD48+ macrophage cluster included those for biological processes associated with alarmins and pro-inflammatory mediators including leukocyte chemotaxis and migration, humoral immune response, and response to lipopolysaccharide (Fig. 3B). These findings support our hypothesis that MERTK+LYVE1high macrophages could restrain inflammatory fibrosis in frozen shoulder, providing a resolving fibrotic niche conducive to restoring homeostasis. Immunostaining of shoulder capsule tissue sections revealed that MerTK+ cells were predominately found in the capsule lining region and were more prevalent in frozen shoulder patient tissues (Fig. 3D & E, P = 0.005). We therefore investigated the cell types predicted to interact with MERTK+LYVE1high and MERTK+LYVE1low macrophage subsets that could restrain inflammatory fibrosis. MerTK, a member of the Tyro-Axl-MERTK (TAM) family of receptor tyrosine kinases is a macrophage receptor that mediates efferocytosis20,21. Engagement of MerTK by apoptotic cells, GAS6 or protein S (PROS1) triggers biochemically distinct responses mediating anti-inflammation and resolution22. PROS1 and GAS6 are ubiquitously expressed by fibroblast populations in the shoulder capsule, particularly APOE+ and CXCL2+ subsets respectively (Fig. 3F). Network Analysis Toolkit for Multicellular Interactions (NATMI) analysis23 generated from comparator and frozen shoulder sub-populations (identified in Fig. 1) predicted specific interactions between MERTK+ macrophages and capsular fibroblasts, including PROS1 > MERTK (Sender cells>Target cells; APOE + >LYVE1high and APOE + >LYVE1low) and GAS6 > MERTK (APOE + >LYVE1high, APOE + >LYVE1low, CXCL2 + >LYVE1high) (Fig. 3G). Further investigation of possible interactions between these TAM ligands and their respective receptors identified GAS6 > AXL fibroblast autocrine circuits (Figure S7A) that were found to be most specific between APOE + > CLIC5+ and CXCL2 + > CLIC5+ clusters of the capsule lining (Figure S7B). Immunostaining confirmed the proximity of MerTK+ macrophages and GAS6+ and PROS1+ fibroblasts in frozen shoulder patient tissues (Fig. 3H). These findings further support the hypothesis that cellular interactions between MERTK+ macrophages and secreted ligands PROS1 and GAS6 from APOE+ & CXCL2+ fibroblasts could provide a resolving fibrotic niche conducive to restoring homeostasis in frozen shoulder.
Fig. 3: Capsular MERTKhigh macrophages have a modulatory phenotype.
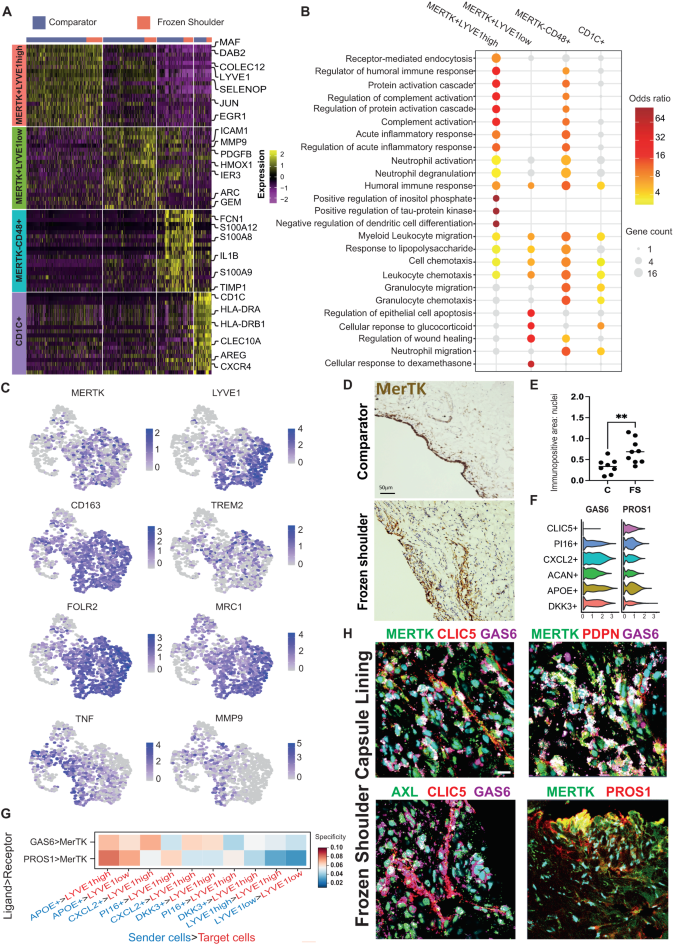
A The heatmap shows the top 20 marker genes for the capsular myeloid cell clusters (two-sided Wilcoxon test, BH adjusted P < 1 × 10−9. B Geneset over-representation analysis of gene ontology (GO) Biological Processes (BP) in the capsular myeloid clusters (one-sided Fisher tests, BH adjusted P < 0.05. C UMAPs show the expression of modulatory macrophage genes including MERTK, LYVE1, CD163, TREM2, FOLR2 and MRC1 in capsular macrophages, localising these markers to MERTK+LYVE1high and MERTK+LYVE1low clusters. D Representative images of 3,3′-diaminobenzidine immunostaining (brown) for MERTK in sections from comparator and frozen shoulder patient tissues, staining is localised to the capsule lining region. Nuclear counterstain is haematoxylin, scale bar=50μm. E Graph shows quantitative analysis of MERTK+ cells in comparator (C, n = 8 donors) and frozen shoulder (FS, n = 9 donors) patient tissues pooled from 2 independent experiments, bars show median values. Statistically significant differences were calculated using a two-sided Mann-Whitney test (P = 0.0055). F Violin plots show expression of MERTK ligands GAS6 and PROS1 in capsular fibroblast sub-populations. G Selected predicted ligand-receptor interactions between sender population (PROS1+ or GAS6+ fibroblasts) and receiver population (MERTK+LYVE1high or MERTK+LYVE1low macrophages) are shown. Predictions were generated from comparator (n = 6 donors) and frozen shoulder (n = 4 donors), sub-populations as in Fig. 1G (NATMI specificity score). H Representative immunofluorescence images of the lining region of frozen shoulder patient tissues, showing the topographical proximity of MERTK with associated ligands GAS6 and PROS1. Fibroblast markers include CLIC5, PDPN, AXL. Scale bar=20μm.
MerTKhigh and MerTKlow macrophages induce divergent responses in capsular fibroblasts from frozen shoulder patients in co-culture
To investigate how capsular macrophages and stromal cells might function to restore homeostasis during frozen shoulder, we explored how the major macrophage subtypes present in the shoulder capsule could influence the phenotype of capsular fibroblasts. To model MerTKhigh and MerTKlow macrophage populations identified in the shoulder capsule, we treated monocyte-derived macrophages (MDMs) with LPS (10 ng/ml)24 or Dexamethasone (1uM) to induce MerTKlow or MerTKhigh phenotypes respectively (validation of respective MerTK phenotypes are shown in Figure S7C & D). We directly co-incubated ex-vivo capsular fibroblasts from frozen shoulder patient tissues in the presence of either MerTKlow or MerTKhigh MDMs for 48 hrs and performed bulk RNAseq on FACS sorted fibroblasts (Figure S7E). We compared the phenotypes of frozen shoulder capsular fibroblasts co-cultured with MerTKlow or MerTKhigh MDMs relative to untreated fibroblasts (Figure S8A). In total, we identified n = 832 genes that showed significant variation in expression across the 3 groups comprising MerTKlow-MDM co-cultured, MerTKhigh-MDM co-cultured capsular fibroblasts, or capsular fibroblasts in isolation (DESeq2, LRT, BH adjusted p < 0.01). These genes clustered into four groups with distinct patterns of expression across the three experimental conditions (Figure S8B), comprising of (i) a set of 345 genes that showed reduced expression in the MerTKhigh-MDM co-cultured fibroblasts relative to untreated and MerTKlow-MDM co-cultured fibroblasts (group 1), (ii) a set of 311 genes which showed higher expression in MerTKhigh-MDM co-cultured fibroblasts (group 2), (iii) a set of 98 genes which showed higher expression in the MerTKlow-MDM co-cultured fibroblasts (group 3) and (iv) a set of 78 genes which showed higher expression in the untreated fibroblasts (Figure S8B with selected examples shown in Fig. 4A). The set of genes downregulated by MerTKhigh MDMs in capsular fibroblasts relative to MerTKlow MDM co-cultured and untreated fibroblasts (group 1) included pro-inflammatory genes (TNFAIP6, PTGES, PTGS2, IL6), chemotactic factors (CXCL3) and matrix metalloproteinases (MMP1). We noted that several of these genes, including IL6, MMP1 and CXCL3 showed higher expression in the fibroblasts co-cultured with MerTKlow-MDMs relative to the untreated fibroblasts (Fig. 4A, Supplementary Data 5). Conversely, MerTKhigh MDMs induced a higher expression of genes (group 2) associated with maintenance and organization of the extracellular matrix (NEXN, GSN, DPT, TIMP4, SPP1), cell adhesion & migration (VIT, EPHB6), cell-matrix interactions (THBS1), immunoregulation (FKBP5) and cell proliferation (TRNP1) relative to MerTKlow-MDM co-cultured and untreated capsular fibroblasts (Fig. 4A, Figure S8B, Supplementary Data 5), suggestive of a more homeostatic response. Co-incubation with MerTKhigh MDMs also down-regulated TGFβ responsive genes including BMP2, BMP4, TGFBR1 and TGFB2 (group 1, Supplementary Data 5). In support of these findings, the set of genes downregulated by co-incubating fibroblasts with MerTKhigh MDMs was over-represented for biological processes including positive regulation of type 2 immune response, prostaglandin biosynthesis and transport, positive regulation of macrophage activation and epithelial to mesenchymal transition and negative regulation of tissue remodelling and apoptotic signalling (group 1, Fig. 4B, Supplementary Data 5). Conversely, genes up-regulated by MerTKhigh MDMs in capsular fibroblasts were over-represented for the biological processes ‘negative regulation of leucocyte degranulation’, ‘negative regulation of IL-1β production’ and those associated with cell-matrix adhesion, phagocytosis recognition, extracellular matrix assembly and disassembly, tissue remodelling and collagen metabolic processes (group 2, Fig. 4B, Supplementary Data 5).
Fig. 4: MerTKlow and MerTKhigh macrophages induce divergent responses in capsular fibroblasts from frozen shoulder patients.
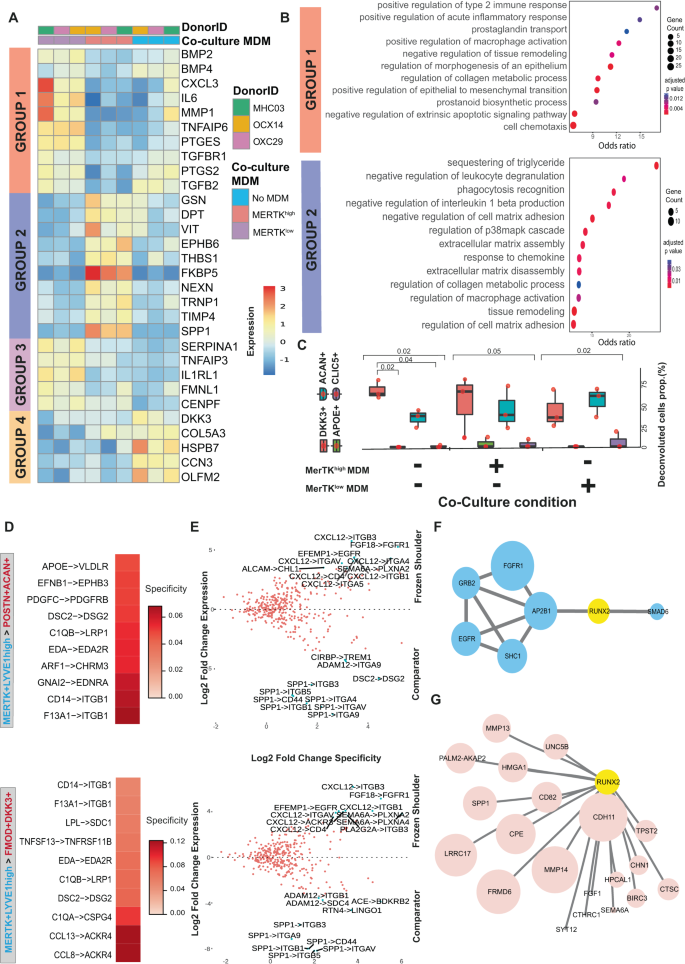
A Heatmap showing selected genes that showed significant variance in expression between capsular fibroblasts from frozen shoulder patients co-incubated with MerTKhigh, MerTKlow MDMs or fibroblasts in isolation (DESeq2, LRT test, BH adjusted P < 0.05). See also Supplementary Fig. 8B. B Dot plot shows GO biological processes over-represented in the sets of genes down-regulated in capsular fibroblasts by incubation with MerTKhigh MDMs (group 1) or up-regulated by incubation with MerTKhigh MDMs (group 2). C The boxplots show the predicted proportions of fibroblast subsets identified in the single-cell analysis (Fig. 1J) present in the untreated, MerTKhigh-MDM co-cultured and MerTKlow-MDM co-cultured fibroblasts (n = 3 frozen shoulder patient donors in 2 independent experiments, bars show median value (median; centre, box limits; lower and upper quartile, whiskers; 1.5 IQR), deconvolution performed with MuSiC). D Selected predicted ligand-receptor interactions between MerTK+LYVE1+ myeloid and DKK3 + FMOD+ or POSTN + ACAN+ fibroblast sub-populations generated from differentially expressed genes from comparator (n = 6 donors) and frozen shoulder (n = 4 donors), sub-populations as in Fig. 1 (NATMI analysis). E The scatter plot shows the expression change of predicted ligand-receptor interactions between MerTK+LYVE1+ myeloid and DKK3 + FMOD+ or POSTN + ACAN+ fibroblast sub-populations in frozen shoulder relative to comparator patient tissues. F Protein-protein network association analysis of receptors (blue) highly expressed in frozen shoulder patient fibroblasts identified a candidate interaction with RUNX2 (analysis performed with IntAct). G Single-cell PySCENIC66 gene regulatory network analysis of fibroblasts (Fig. 1J, all clusters) identified a connection between the expression of RUNX2 and matrix associated genes including CDH11, MMP14, MMP13 and SPP1 (pink) in POSTN + ACAN+ fibroblasts.
We next assessed if co-culturing MerTKlow or MerTKhigh MDMs with capsular fibroblasts influenced the composition of fibroblast sub-populations in vitro. Deconvolution of bulk RNAseq data relative to fibroblast populations in Fig. 1J predicted that fibroblasts in the resting state (in the absence of MDMs) were comprised of DKK3 + FMOD+ (median 61%) and POSTN + ACAN+ (median 35%) sub-populations (Fig. 4C). Co-culture with MerTKhigh MDMs did not induce a significant shift in the predicted composition. In contrast, co-culture with MerTKlow MDMs increased the predicted percentage of POSTN + ACAN+ (med 58%) cells and decreased the predicted percentage of DKK3 + FMOD+ (med 33%) cells. These findings suggest that under homeostatic and resolving inflammatory milieu, DKK3 + FMOD+ and POSTN + ACAN+ subsets are the predominant capsular fibroblasts sub-populations in vitro, and that MerTKlow MDMs can promote development of a POSTN + ACAN+ fibroblast phenotype at the expense of a DKK3 + FMOD+ phenotype. Immunocytochemistry confirmed expression of DKK3 and POSTN proteins in incubations of frozen shoulder capsular fibroblasts co-cultured with either MerTKhigh or MerTKlow MDMs (Figure S7F, isotype controls shown in Figure S7G).
We applied NATMI cell-cell communication analysis to identify candidate signaling factors in MERTK+LYVE1high macrophages for the induction of cellular and molecular pathways regulating extracellular matrix organization and structure in the DKK3 + FMOD+ or POSTN + ACAN+ fibroblast populations. Predicted interactions between these cell types were generated from comparator and frozen shoulder sub-populations (as per Fig. 1), identifying ligand-receptor pairs including F13A1 > ITGB1, CD14 > ITGB1, C1QB > LRP1, DCS2 > DGS2, EFNB1 > EPHB3, APOE > VLDLR, CCL8 > ACKR4, CCL13 > ACKR4 and LPL > SDC1 (Fig. 4D). Frozen shoulder patient tissues showed higher expression of predicted ligand-receptor pairs between the MERTK+LYVE1high macrophages and DKK3 + FMOD+ or POSTN + ACAN+ fibroblast populations that included CXCL12 > ITGB3, FGF18 > FGFR1, EFEMP1 > EGFR and CXCL12 > ITGB1 (Fig. 4E). In contrast, we noted that comparator tissues showed a higher expression of predicted interactions involving SPP1 expression by the MERTK+LYVE1high macrophages (Fig. 4E). Protein-protein association analysis with IntAct25 identified a possible link between the receptors highly expressed in frozen shoulder patient tissues and the transcription factor RUNX2 (Fig. 4F). Separately, single-cell gene-regulatory network analysis of POSTN + ACAN+ fibroblasts revealed a correlation between the expression of RUNX2 and genes implicated in remodelling and organisation of the extracellular matrix including CDH11, MMP14, MMP13 and SPP1 (Fig. 4G). Relative to comparator tissues, frozen shoulder patient tissues showed a lower expression of predicted ligand-receptor pairs involving expression of the pro-inflammatory lymphotoxin-beta (LTB) cytokine by MERTK+LYVE1high macrophages and LTB receptors by DKK3 + FMOD+ or POSTN + ACAN+ fibroblasts (including LTB > CD40, LTB > LTBR and LTB > TNFRSF1A, Figure S8C). Collectively these findings suggest that cellular interactions between MERTK+ macrophages and DKK3 + FMOD+ and POSTN + ACAN+ fibroblasts may play a role in the resolution of frozen shoulder by inducing integrin-mediated remodelling of the fibrotic extracellular matrix and restraining the inflammatory phenotype of capsular fibroblasts.
Cross-tissue comparison of MERTK+ macrophages in musculoskeletal tissues
Having identified a potential modulatory phenotype for MERTK+ macrophages in patient shoulder capsule tissues, we next compared capsular MERTK+ macrophages with a published dataset of MERTK+ synovial tissue macrophages (STM) isolated from rheumatoid arthritis (RA) patient cohorts24. MERTK + STM sub-populations were associated with RA disease remission, showed enrichment for negative regulators of inflammation and induced repair responses in synovial fibroblasts in vitro24. The transcriptome correlation distance between shoulder capsule and knee synovial MERTK+ and MERTK–CD48+ myeloid sub-populations is shown in Fig. 5A. MERTK+LYVE1high macrophage clusters were common to both tissue types, the equivalent MERTK+ cluster in knee RA synovial tissues is annotated FOLR2highLYVE1 + 24 (Fig. 5A). We applied a transfer-learning approach26 to annotate the STM populations (Fig. 5B) with the cluster labels from our shoulder capsule dataset (Fig. 1G). The MERTK+LYVE1high macrophage cluster in the shoulder capsule mapped to TREM2low, TREM2high, FOLR2highLYVE1+ and FOLR2 + ID2 + STM sub-populations (Fig. 5C, reverse label transfer shown in Figure S8D).
Fig. 5: Comparison of MERTKhigh macrophage clusters in shoulder capsule and knee RA synovial tissues.
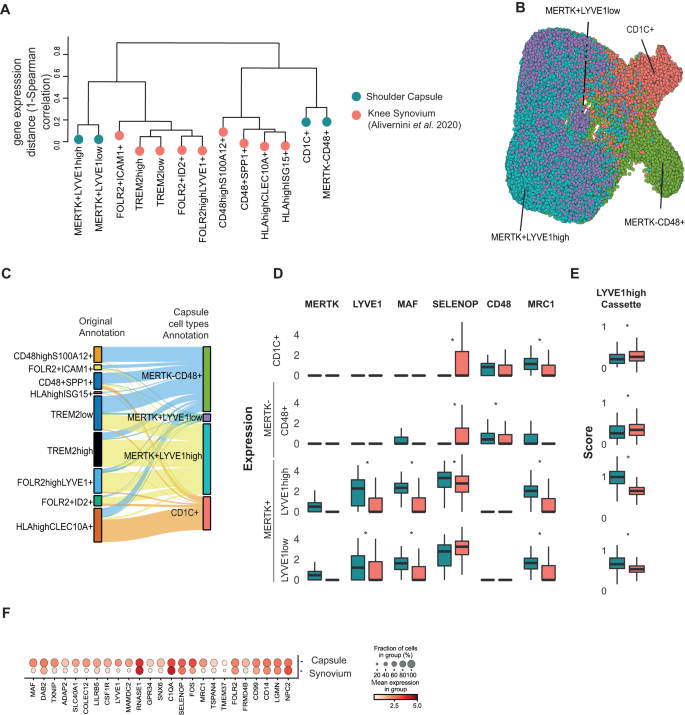
A Dendrogram shows the transcriptomic Spearman correlation distance between human myeloid populations in comparator and frozen shoulder capsule tissues (blue) relative to knee RA synovial tissue macrophages (STMs, red) from Alivernini et al. (2020). In STMs, the MERTK + LYVE1+ cluster is annotated FOLR2highLYVE1+. B UMAP shows the Alivernini et al. (2020) STM cells with labels transferred from the myeloid clusters identified in adult shoulder capsule tissues (Fig. 1G) (scArches analysis) C The Sankey plot shows the mapping between the original STM sub-populations clusters identified by Alivernini et al. (2020) (left) and the transferred myeloid labels from the adult shoulder capsule tissues (MERTK–CD48+, MERTK+LYVE1low, MERTK+LYVE1high, CD1C+) (right). D The box plots show the normalized expression of myeloid genes including MAF, LYVE1, MERTK, SELENOP, CD48 and MRC1 in the shoulder capsule myeloid clusters (blue, as per Fig. 1G, CD1C+ cluster n = 7; other cell types n = 8) and the corresponding STM subsets (red, as predicted by label transfer, all cell types n = 10). Bars show median value, *P < 0.05 (Wald test). E Box plots (right) show cassette scores for marker genes of the MERTK+LYVE1high cluster in the shoulder capsule myeloid clusters (Fig. 1G, CD1C+ cluster n = 7; other cell types n = 8) and corresponding knee STM subsets (as predicted by label transfer, all cell types n = 10). Bars show median value, *P < 0.05 (Wald test). Box plots in D and E show median as centre, lower and upper quartiles as box limits, and whiskers with the length of 1.5 IQR. F Dot plot shows average expression of top 25 MERTK+LYVE1high macrophage cluster marker genes in the shoulder capsule MERTK+LYVE1high macrophages relative to their corresponding cells in the Alivernini et al. (2020) STMs (as predicted by label transfer). Box plots in D and E show median as centre, lower and upper quartiles as box limits, and whiskers with the length of 1.5 IQR.
Using transferred labels, we investigated the relative expression of myeloid target genes of interest including MAF, LYVE1, MERTK, SELENOP, CD48 and MRC1 in the shoulder capsule clusters and their corresponding (′) STMs subsets. Within the MERTK+LYVE1high subsets, expression of genes implicated in restoring tissue homeostasis including LYVE1, MRC1 and MAF was higher in shoulder capsule cells relative to the corresponding STMs subset (Fig. 5D, DESeq2 analysis, Wald Test, BH adjusted P < 0.05). To investigate whether these differences were representative of the broader MERTK+LYVE1high phenotype we used a gene-cassette comprised of n = 224 shoulder capsule cluster marker genes for this population. A lower expression of this gene cassette was observed in the predicted knee synovium MERTK+LYVE1high′ (Wald test, BH adjusted P = 1.31 × 10−199) and MERTK+LYVE1low′ (BH adjusted P = 5.75 × 10−37) populations than was seen in the corresponding shoulder capsule clusters (Fig. 5E). We confirmed that this pattern was recapitulated after performing a reverse label transfer to identify a shoulder capsule FOLR2highLYVE1+′ cluster analogue (Figure S8D, E). Inspection of the expression of the top 25 markers of MERTK+LYVE1high shoulder capsule macrophages (Fig. 1) confirmed that while this capsule population and its predicted MERTK+LYVE1high synovial counterpart had similar levels of expression of several marker genes including C1QA, SELENOP, FOLR2 and CD14, the expression of others, including LYVE1, MAF and MRC1 was less pronounced in the MERTK+LYVE1high′ STM (Fig. 5F). Collectively, these findings show that while MERTK+ macrophages in the shoulder capsule and RA synovial patient tissues share a common gene expression cassette, MERTK+ macrophages in shoulder capsule patient tissues had a higher expression of genes implicated in restoring tissue homeostasis. To further understand the role of MERTK+ macrophages in frozen shoulder and RA, we compared gene expression changes in MERTK+ macrophages in these two diseases between patients and respective healthy controls. Overall, we observed a correlation of 0.21 (Wilcoxon test, P = 2.42 × 10−5) in the transcriptional changes within the MERTK+ macrophages between the two diseases (Figure S8F). To pin-point genes that were regulated in different directions in disease in the two tissues we performed an interaction analysis. This identified 6 statistically significant genes that showed different directions of regulation in frozen shoulder vs RA. Relative to their healthy counterpart, we observed AQP1, CLEC11A, CRIP2, EGR1 and SEMA4A being upregulated in diseased knee RA synovial tissues and downregulated in diseased frozen shoulder tissues (Figure S8G). Conversely, BID was upregulated in diseased frozen shoulder tissues and downregulated in diseased RA knee tissues relative to healthy comparators (Figure S8G, Supplementary Data 3). Despite these relatively minor differences, the overall similarity of the MERTK+ macrophage phenotype in frozen shoulder and RA supports the concept that this subset of macrophages may also be important for the resolution of frozen shoulder.
The developing shoulder joint provides a cellular template for resolution
Ontological processes can be recapitulated as part of inflammatory disease pathology27. To explore if the cell types implicated in the resolution of inflammatory fibrosis in frozen shoulder are present during the development of the human foetal shoulder joint, we generated an atlas of human developmental shoulder tissues at 12, 15 and 17 post-conception weeks (pcw), (Supplementary Data 1). Following integration, clustering of 12,661 cells revealed 9 major populations, each represented in cells from 12, 15 and 17 pcw developmental stages (Fig. 6A, Figure S9A & Supplementary Data 6). The major cell types identified in our dataset include stromal fibroblasts, myeloid, progenitor and differentiated T cells and B cells, vascular endothelial cells, mural and cycling cells (Fig. 6A). Lymphoid populations were identified in foetal shoulder tissues from 15 and 17pcw developmental stages. These included CD3 + CD8+ T cells (GZMB + NKG7+), CD3 + CD4+ T cells (IL7R+) (Figure S9B), differentiated MS4A1 + B cells and three CD34+ B-cell progenitor clusters (CD38 + IGLC1+, MDM2 + RAG1+ and MKI67 + TOP2A+, Figure S9C). Immunostaining of developmental shoulder capsule tissues revealed CD19+ cells reside within CD31+ blood vessels, suggesting identified B cells as possible blood contaminants (Figure S9D). Clustering of foetal CD68 + CD14+ cells revealed 7 distinct populations including MERTK+LYVE1highMφ, MERTK + LYVE1highTIMD4highMφ, MERTKlowS100A12 + Mφ, cycling MKI67 + TOP2A + Mφ, MYL4+ myeloid progenitors, CD1C+ monocyte-derived dendritic cells and CAECAM8+ neutrophils (Fig. 6B, Figure S9E, Supplementary Data 6).
Fig. 6: The developing shoulder capsule informs a template for resolution.
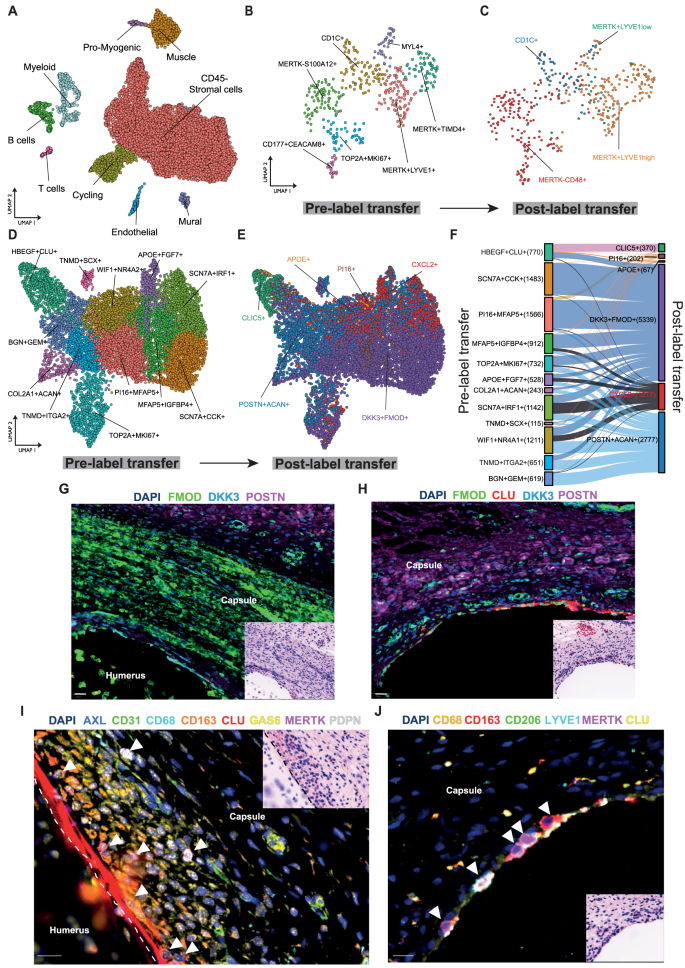
scRNAseq was performed on 12, 15 and 17 pcw developing human shoulder joint tissues. A UMAP shows the major cell types comprising the developing foetal shoulder joint (resolution = 0.9). B UMAP (resolution = 0.6) shows identified myeloid populations in the developing shoulder joint (12, 15 and 17 pcw). Sub-clustering of myeloid populations revealed 7 distinct clusters including MERTK + LYVE1+ and MERTK+TIMD4+ populations. C The plot shows the same foetal cell UMAP with labels transferred from the adult shoulder capsule myeloid cells (Fig. 1G) (scArches analysis). D UMAP (resolution = 0.7) showing CD45–COL1A1+ stromal cell clusters in the developing shoulder joint (12, 15 and 17 pcw). E The plot shows the same UMAP of developing shoulder stromal cells with labels transferred from the adult clusters (Fig. 1J) (scArches analysis). F The Sankey plot shows the mapping between the clusters identified in the developing shoulder stromal cells (D) (left) and the transferred labels from the adult fibroblast populations (right). G–J Representative Cell DIVE and respective H&E stained images of histological sections of foetal shoulder joint at 17 post conception weeks (pcw) development stage. G, H Sections stained for a panel of markers to identify fibroblasts in the shoulder capsule during development (FMOD, DKK3, POSTN, CLU). DAPI nuclear counterstain is dark blue, scale bar = 20 μm. I High magnification image showing the cell types identified in foetal shoulder capsule including macrophages (CD68, CD163, MerTK) and fibroblasts (PDPN, CLU, AXL, GAS6) in the capsule lining region at 17 pcw. Scale bar = 20 μm. J High magnification image of the foetal shoulder capsule (17 pcw) showing immunostaining for macrophage markers MerTK, LYVE1, CD206 and CD163 in the capsule lining region. DAPI nuclear counterstain is dark blue, white arrows mark MerTK+ macrophages. Scale bar = 20 μm.
Having identified interactions between MERTK+LYVE1high macrophages and matrix associated DKK3 + FMOD+ and POSTN + ACAN+ fibroblasts induce matrix remodelling in cells from adult tissues, we investigated if correlates of these adult cell populations are present during foetal development. To do so, we transferred the cluster labels from the adult shoulder capsule myeloid clusters (as in Fig. 1G) onto foetal myeloid cells. The adult MERTK+LYVE1high cluster mapped to TOP2A + MKI67+, MERTK + TIMD4+, MYL4+, MERTK + LYVE1+ and CD1C+ foetal myeloid sub-populations (Figure S9F). This analysis suggested that the MERTK+LYVE1high macrophage phenotype is common to both adult and foetal shoulder tissues (Fig. 6B).
Analysis of foetal stromal cells identified fibroblast clusters including PI16 + MFAP5+, SCN7A + CCK+, WIF1 + NR4A2 + SCN7A + IRF1+, MFAP5 + IGFBP4+, TNMD + ITGA2, BGN + GEM+, APOE + FGF7+, MKI67 + TOP2A+ and HBEGF + CLU+ (lining layer) sub-populations, COL2A1 + ACAN+ chondrocyte and TNMD + SCX+ tendon cell clusters (Fig. 6D, Figure S9G, Supplementary Data 6). Transfer of labels from adult fibroblast clusters (Fig. 1J) onto respective foetal fibroblasts suggested that similar DKK3 + FMOD+ and POSTN + ACAN+ fibroblast populations may be present in foetal shoulder tissues (Fig. 6E). The adult DKK3 + FMOD+ cluster maps to all foetal fibroblast sub-populations, the adult POSTN + ACAN+ cluster maps to all foetal fibroblast sub-populations except for SCN7A + CCK+ and HBEGF + CLU clusters (Fig. 6F). Feature plots showing expression of markers for major adult fibroblast populations (DKK3 + FMOD+ and POSTN + ACAN+) in foetal CD45– cells are shown in Figure S9H.
Finally, we spatially mapped myeloid and fibroblast populations implicated in the resolution of frozen shoulder onto foetal shoulder joint tissues to compare the respective topographical niches of these cells with adult tissues. Representative images of foetal shoulder joints are shown in Figure S10A & B, highlighting the anatomy of the shoulder joint capsule at 14 and 17pcw developmental stages. Immunostaining confirmed the presence of major cell types including fibroblasts (CD34, CD90), macrophages (CD68), T cells (CD4) and vascular endothelium (CD31, CD146) within the developing shoulder capsule (Figure S10C). CLU+ fibroblasts delineated the border between capsular fibroblasts and adjacent proximal humerus, NOV+ fibroblasts resided in the deeper sub-lining region (Figure S10 D & E). DKK3 + FMOD+ and POSTN+ fibroblasts were identified in the developing shoulder capsule (Fig. 6G & H). MERTK + LYVE1 + MRC1+ macrophages localised to the lining region of the developing shoulder capsule adjacent to CLU+ and GAS6+ fibroblasts (Fig. 6I & J, Figure S10F). Respective images for isotype control staining are shown in Figure S10G & H. Collectively these scRNAseq and spatial findings support an embryonic origin for MERTK+ macrophages and DKK3 + FMOD+ and POSTN+ fibroblasts in the shoulder capsule. This data supports the conclusion that the cell types implicated in the resolution of adult shoulder fibrosis are present during foetal development.
Discussion
Frozen shoulder is a naturally occurring chronic inflammatory fibrotic human disease that is uniquely self-limiting over time. Analysis of well-phenotyped tissues collected from male and female patients during the advanced resolving disease stage provides an opportunity to identify the cell types and molecules implicated in fibrosis resolution. Using integrated scRNA-seq, spatial validation, cross-tissue comparison, and in vitro mechanistic experiments we provide new insights into the functional biology of this resolving inflammatory fibrotic niche. Systematic analysis of the cell types comprising the adult shoulder capsule during resting and diseased states identified distinct subsets of fibroblasts, T cells and macrophages occupying discrete microanatomical niches. Capsular fibroblasts share common features with those previously described in synovial tissues16,18,19,24. We identified two matrix-associated capsular fibroblast populations (DKK3 + FMOD+ and POSTN + ACAN+), two chemoattractant clusters (CXCL12 + APOE+ and CXCL2 + ARC+), vascular interacting (PI16 + MFAP5+) and a lining layer cluster (CLIC5 + HBEGF+). Frozen shoulder patient tissues showed increased cellularity of lining and sub-lining regions relative to comparator patient tissues and highly expressed POSTN and CTHRC1 in the capsule sub-lining. Frozen shoulder patient tissues were enriched for T cells which also resided in the capsule sub-lining. CD4 + CD127+ T cells expressed Th17-associated genes including CCR6, RORA and AHR as well as genes associated with a tissue repair phenotype28,29,30. This CD4+ subset was distinct from CD4+ clusters identified in patient RA synovial tissues31. CD8+ T cell and NK subsets expressed granzyme, granulysin and perforin molecules akin to the SCT5 subset identified in knee synovial tissues from RA patients31 and other inflamed human tissues32. Future studies are required to determine if these CD8+ cells contribute to resolution of frozen shoulder by facilitating the clearance of senescent cells.
토론
동결견은 시간이 지남에 따라
자연적으로 자가 제한되는 만성 염증성 섬유화 인간 질환이다.
진행성 해소 단계의 남녀 환자에서 채취한 잘 표현형화된 조직 분석은
섬유화 해소와 관련된 세포 유형 및 분자를 식별할 기회를 제공한다.
통합 단일세포 RNA-seq, 공간적 검증, 조직 간 비교 및 체외 기전 실험을 통해
우리는 이 해결 중인 염증성 섬유화 틈새의 기능적 생물학에 대한
새로운 통찰력을 제공한다.
휴지 상태와 질환 상태에서 성인 어깨 관절낭을 구성하는 세포 유형에 대한 체계적 분석은
서로 다른 미세해부학적 틈새를 점유하는
섬유아세포, T 세포 및 대식세포의 별개의 하위 집단을 확인했다.
낭막 섬유아세포는
관절막 조직에서 이전에 보고된 섬유아세포와 공통된 특징을 공유한다16,18,19,24.
우리는 두 가지 기질 관련 캡슐 섬유아세포 집단(DKK3⁺ FMOD⁺ 및 POSTN⁺ ACAN⁺), 두 가지 화학유인인자 클러스터(CXCL12⁺ APOE⁺ 및 CXCL2⁺ ARC⁺), 혈관 상호작용 집단(PI16⁺ MFAP5⁺), 그리고 내벽층 클러스터(CLIC5⁺ HBEGF⁺)를 확인했습니다. 동결견 환자 조직은 대조군 환자 조직에 비해 내피 및 하부 내피 영역의 세포 밀도가 증가했으며, 캡슐 하부 내피에서 POSTN 및 CTHRC1을 고도로 발현했습니다. 동결견 환자 조직은 캡슐 하부 내피에도 존재하는 T 세포가 풍부했습니다. CD4⁺ CD127⁺ T 세포는 CCR6, RORA 및 AHR을 포함한 Th17 관련 유전자와 조직 복구 표현형과 관련된 유전자들을 발현했다28,29,30. 이 CD4+ 하위 집합은 류마티스 관절염(RA) 환자 활막 조직에서 확인된 CD4+ 클러스터와 구별되었습니다31.
CD8+ T 세포 및 NK 하위 집합은
RA 환자 무릎 활막 조직31 및 기타 염증성 인간 조직32에서 확인된
SCT5 하위 집합과 유사하게 그랜자임, 그라눌리신 및 퍼포린 분자를 발현했습니다.
이러한 CD8+ 세포가
노화 세포 제거를 촉진하여 동결견 해결에 기여하는지 확인하기 위해서는
추가 연구가 필요합니다.
MERTKlowCD48+ macrophages from frozen shoulder patients highly expressed IL1A and IL1B although this inflammatory macrophage cluster showed reduced SPP1 expression relative to tissues from C-19 and RA patients33. While it is plausible that this population of cells may play a targetable role in disease pathogenesis, future studies of patient tissues collected during earlier-stage frozen shoulder are essential to confirm if these inflammatory mediators are also active during early-stage disease. A population of MERTK + LYVE1 + MRC1+ macrophages resident in the capsule lining was enriched for biological processes concerned with modulating inflammation including regulation of humoral immune response, complement activation, inflammatory response, and receptor-mediated endocytosis. These findings support the hypothesis that MERTK+ macrophages might restrain inflammatory fibrosis, providing a resolving fibrotic niche conducive to restoring homeostasis in frozen shoulder, as observed at other sites of tissue repair2,34,35,36.
동결견 환자의 MERTKlowCD48+ 대식세포는
IL1A 및 IL1B를 고도로 발현했으나,
이 염증성 대식세포 군집은 C-19 및 RA 환자 조직에 비해
SPP1 발현이 감소된 것으로 나타났다33.
이 세포 집단이 질병 발병 기전에서 표적 가능한 역할을 할 가능성이 있지만,
이러한 염증 매개체들이 초기 단계 질환에서도 활성 상태인지 확인하기 위해서는
동결견 초기 단계에서 채취한 환자 조직에 대한 향후 연구가 필수적이다.
캡슐 내막에 상주하는 MERTK+ LYVE1+ MRC1+ 대식세포 집단은
체액성 면역 반응 조절, 보체 활성화, 염증 반응, 수용체 매개 세포 내 흡수 등
염증 조절과 관련된 생물학적 과정에 풍부하게 관여하고 있었다.
이러한 결과는
MERTK+ 대식세포가 염증성 섬유화를 억제하여,
다른 조직 복구 부위에서 관찰된 바와 같이 동결견에서 항상성 회복에 유리한
섬유화 소멸 환경을 제공할 수 있다는 가설을 뒷받침한다2,34,35,36.
Postulating that capsular MERTK+ macrophages aid resolution of frozen shoulder, we investigated how cellular crosstalk between MERTK+ macrophages and capsular fibroblasts from frozen shoulder patients could underpin fibrosis resolution. Co-incubation of capsular fibroblasts from frozen shoulder patients with MerTKlow MDMs induced an inflammatory fibroblast phenotype and biological pathways converging on chemotaxis and prostanoid biosynthesis. Conversely, co-incubation with MerTKhigh MDMs negatively regulated leucocyte degranulation and IL-1β production pathways and upregulated genes and pathways associated with maintenance and organisation of the extracellular matrix and down-regulated TGFβ responsive genes including BMP2, TGFBR1 and TGFB2 relative to frozen shoulder capsular fibroblasts in isolation. Given that TGFβ induces the myCAF phenotype in cancer-associated fibroblasts37, downregulation of TGFβ signalling could modulate the desmoplastic phenotype of capsular fibroblasts during frozen shoulder.
관절낭 내 MERTK+ 대식세포가 동결견의 해결을 돕는다는 가설을 바탕으로, 동결견 환자의 관절낭 섬유아세포와 MERTK+ 대식세포 간의 세포 간 교신이 어떻게 섬유화 해결을 뒷받침할 수 있는지 조사하였다. 동결견 환자의 관절낭 섬유아세포를 MerTKlow MDMs와 공동 배양했을 때, 염증성 섬유아세포 표현형과 화학유인 및 프로스타노이드 생합성에 수렴하는 생물학적 경로가 유도되었다. 반대로, MerTKhigh MDM과의 공동 배양은 백혈구 탈과립 및 IL-1β 생성 경로를 음성 조절하고, 세포외 기질의 유지 및 조직화와 관련된 유전자 및 경로를 상향 조절하며, 단독으로 배양된 동결견 관절낭 섬유아세포에 비해 BMP2, TGFBR1 및 TGFB2를 포함한 TGFβ 반응성 유전자를 하향 조절했습니다. TGFβ가 암 관련 섬유아세포에서 myCAF 표현형을 유도한다는 점을 고려할 때37, TGFβ 신호전달의 하향조절은 동결견 동안 관절낭 섬유아세포의 섬유성 변성 표현형을 조절할 수 있다.
A recent study reported expansion of the DKK3+ synovial fibroblasts in treatment-refractory RA patients38, however the precise function of this fibroblast population remains to be established39,40. Interestingly, DKK3 + FMOD+ fibroblasts in the shoulder capsule highly expressed inflammation-resolving receptor CMKLR1, which we previously identified in advanced-stage frozen shoulder patient tissues14. DKK3 + FMOD+ and POSTN + ACAN+ fibroblasts were the predominant populations isolated from patient shoulder capsule tissues utilised in micro-culture experiments with MerTKhigh and MerTKlow macrophages. Predicted interactions between these cell populations revealed enrichment for ligand-receptor pairs including F13A1 > ITGB1, CD14 > ITGB1, C1QB > LRP1, DCS2 > DGS2, EFNB1 > EPHB3, APOE > VLDLR, CCL8 > ACKR4, CCL13 > ACKR4 and LPL > SDC1. Notably, frozen shoulder patient tissues showed enrichment for ligand-receptor pairs including CXCL12 > ITGB3, FGF18 > FGFR1, EFEMP1 > EGFR and CXCL12 > ITGB1. ITGB1, ITGB3, EPHB3 and SDC1 are matrix-associated molecules. Fibroblast expression of integrins including ITGB1 is required to support tissue repair and remodelling41. Eph receptors and their ligands are essential to the development and function of neuromuscular junctions and regulate actin cytoskeletal dynamics42,43. Syndecans including SDC1 mediate cytoskeletal organisation and remodelling of the extracellular matrix in cancer44. LRP1 promotes phagocytic activity, facilitating clearance of apoptotic cells45 required for return towards tissue homeostasis. Interestingly, frozen shoulder patient tissues showed depletion for ligand-receptor pairs driving inflammatory processes including LTB > CD40, LTB > LTBR and LTB > TNFRSF1A. Protein-protein network association analysis of receptors highly expressed in frozen shoulder patient fibroblasts identified a possible role for the RUNX2 transcription factor in regulating the expression of matrix-associated genes including CDH11, MMP14, MMP13 in these cells. Collectively, these experiments using patient-derived cells inform interactions between MERTK+ macrophages and matrix-associated DKK3 + FMOD+ and POSTN + ACAN+ fibroblasts play a role in the resolution of frozen shoulder by inducing integrin mediated remodelling of the fibrotic extracellular matrix and restraining the inflammatory phenotype of capsular fibroblasts, suggesting a pro-resolving role for these cell populations in fibrosis resolution.
최근 연구에서는 치료 불응성 류마티스 관절염 환자에서 DKK3+ 활막 섬유아세포의 증식이 보고되었으나38, 이 섬유아세포 집단의 정확한 기능은 아직 규명되지 않았다39,40. 흥미롭게도, 어깨 관절낭 내 DKK3+ FMOD+ 섬유아세포는 염증 해소 수용체인 CMKLR1을 고도로 발현했는데, 이는 우리가 이전에 진행성 동결견 환자 조직에서 확인한 바 있다14. DKK3+ FMOD+ 및 POSTN+ ACAN+ 섬유아세포는 MerTKhigh 및 MerTKlow 대식세포와의 미세 배양 실험에 사용된 환자 어깨 관절낭 조직에서 분리된 주요 집단이었다. 이러한 세포 집단 간의 예측 상호작용 분석 결과, F13A1 > ITGB1, CD14 > ITGB1, C1QB > LRP1, DCS2 > DGS2, EFNB1 > EPHB3, APOE > VLDLR, CCL8 > ACKR4, CCL13 > ACKR4 및 LPL > SDC1과 같은 리간드-수용체 쌍의 풍부함을 보여주었다. 특히 동결견 환자 조직에서는 CXCL12 > ITGB3, FGF18 > FGFR1, EFEMP1 > EGFR 및 CXCL12 > ITGB1과 같은 리간드-수용체 쌍의 풍부함이 관찰되었다. ITGB1, ITGB3, EPHB3 및 SDC1은 매트릭스 관련 분자입니다. ITGB1을 포함한 인테그린의 섬유아세포 발현은 조직 복구 및 재구성을 지원하기 위해 필요합니다41. Eph 수용체와 그 리간드는 신경근 접합부의 발달과 기능에 필수적이며 액틴 세포골격 동역학을 조절한다42,43. SDC1을 포함한 신데칸은 암에서 세포골격 조직화와 세포외 기질의 재구성을 매개한다44. LRP1은 식세포 활성을 촉진하여 조직 항상성 회복에 필요한 세포 사멸 세포의 제거를 용이하게 합니다45. 흥미롭게도, 오십견 환자 조직에서는 LTB>CD40, LTB>LTBR 및 LTB>TNFRSF1A를 포함한 염증 과정을 유도하는 리간드-수용체 쌍의 고갈이 관찰되었습니다. 오십견 환자 섬유아세포에서 고도로 발현되는 수용체에 대한 단백질-단백질 네트워크 연관성 분석을 통해, RUNX2 전사 인자가 CDH11, MMP14, MMP13 등의 매트릭스 관련 유전자 발현 조절에 RUNX2 전사인자가 관여할 가능성을 제시하였다. 종합적으로, 환자 유래 세포를 이용한 이러한 실험들은 MERTK+ 대식세포와 매트릭스 관련 DKK3+ FMOD+ 및 POSTN+ ACAN+ 섬유아세포 간의 상호작용이 섬유화성 세포외기질의 인테그린 매개 재구성을 유도하고 관절낭 섬유아세포의 염증성 표현형을 억제함으로써 동결견의 해결에 기여함을 보여주며, 이러한 세포 집단이 섬유화 해결에 해결 촉진적 역할을 수행함을 시사한다.
Synovial tissue macrophages (STM) from RA patients are comprised of distinct subsets, with patients in disease remission showing enrichment for MERTK+ clusters including TREM2high and FOLR2highLYVE1+ macrophage sub-populations24. We compared MERTK+ macrophages in the adult shoulder capsule with knee RA STMs, identifying a common gene expression cassette between these tissue types. The overall similarity of the transcriptional signature of the adult shoulder capsule MERTK+ macrophages with remission associated STM populations supports a possible role for this myeloid population in also restoring homeostasis in frozen shoulder patient tissues. Intriguingly, we noted higher expression of the inflammation regulating genes LYVE1, MRC1 and MAF in the frozen shoulder MERTK+ macrophages than was detected on their predicted STM counterparts. This suggests that these cells may have had a more potent resolving phenotype in the shoulder capsule samples in keeping with the unusually self-limiting nature of frozen shoulder.
류마티스 관절염(RA) 환자의 활막 조직 대식세포(STM)는 서로 다른 하위 집합으로 구성되며, 질병 관해 상태의 환자는 TREM2high 및 FOLR2highLYVE1+ 대식세포 하위 집단을 포함한 MERTK+ 클러스터가 풍부하게 나타난다24. 성인 어깨 관절낭 내 MERTK+ 대식세포를 무릎 RA STM과 비교하여, 이들 조직 유형 간 공통 유전자 발현 카세트를 확인하였다. 성인 어깨 관절낭 MERTK+ 대식세포의 전사 시그니처가 관해 관련 STM 집단과 전반적으로 유사하다는 점은, 이 골수성 집단이 동결견 환자 조직에서도 항상성 회복에 기여할 가능성을 시사한다. 흥미롭게도, 동결견 MERTK+ 대식세포에서 염증 조절 유전자 LYVE1, MRC1 및 MAF의 발현이 예측된 STM 대조군보다 높게 관찰되었다. 이는 동결견의 특이한 자가제한적 특성에 부합하여, 이들 세포가 어깨 관절낭 샘플에서 더 강력한 해결형 표현형을 가졌을 가능성을 시사한다.
Murine studies highlight perivascular MERTK+LYVE1high macrophages maintain tissue homeostasis and prevent arterial stiffness and collagen deposition via MMP9-dependent proteolysis46. Depletion of LYVE1highMHCIIlow monocyte-derived resident tissue macrophages in a murine lung fibrosis model exacerbated vessel permeability, immune cell infiltration, and collagen deposition, demonstrating their critical role in restraining inflammation and fibrosis47. In synovial tissues, MERTK + STM from remission RA patients were potent producers of inflammation-resolving lipids and induced repair responses in cultured synovial fibroblasts24. Collectively, these findings further support the ability of MERTK+ macrophages to restrain inflammation and support homeostasis across multiple sub-tissular niches. Using microcultures of patient-derived cells, Alivernini et al. identified that MERTK + STM induced synovial repair responses in synovial fibroblasts from RA patients during disease remission24. Our functional experiments revealed that MerTKhigh MDMs restrained the inflammatory phenotype of frozen shoulder capsular fibroblasts and upregulated genes and pathways implicated in extracellular matrix organisation and remodelling which could also affect fibrosis resolution. Therefore, tissue-specific contexts could dictate the nature of this homeostatic response.
생쥐 연구에 따르면 혈관 주위 MERTK+LYVE1high 대식세포는 MMP9 의존성 단백질 분해를 통해 조직 항상성을 유지하고 동맥 경직 및 콜라겐 침착을 방지한다46. 생쥐 폐 섬유증 모델에서 LYVE1highMHCIIlow 단핵구 유래 상주 조직 대식세포를 제거하면 혈관 투과성, 면역 세포 침윤 및 콜라겐 침착이 악화되어 염증과 섬유증을 억제하는 데 있어 이들의 중요한 역할을 입증하였다47. 활막 조직에서, 관해기 류마티스 관절염(RA) 환자의 MERTK+ STM은 염증 해소 지질의 강력한 생성체였으며 배양된 활막 섬유아세포에서 수복 반응을 유도했습니다24. 종합적으로, 이러한 결과들은 MERTK+ 대식세포가 다양한 하위 조직 틈새에서 염증을 억제하고 항상성을 유지하는 능력을 추가로 뒷받침합니다. 환자 유래 세포의 미세 배양을 이용한 Alivernini 등의 연구는 MERTK+ STM이 RA 환자의 관절막 섬유아세포에서 질병 관해 기간 동안 관절막 회복 반응을 유도함을 확인했다24. 우리의 기능적 실험은 MerTKhigh MDM이 동결견 관절낭 섬유아세포의 염증성 표현형을 억제하고, 섬유증 해결에도 영향을 미칠 수 있는 세포외 기질 조직 및 재구성과 관련된 유전자 및 경로를 상향 조절함을 밝혔다. 따라서 조직 특이적 맥락이 이러한 항상성 반응의 본질을 결정할 수 있다.
Having identified that crosstalk between MERTK+ macrophages and matrix associated fibroblasts underpin matrix remodelling in frozen shoulder, we investigated if the cell types implicated in the resolution of adult fibrotic capsular disease are present during foetal development. Single-cell analysis of human foetal shoulder tissues from 12–17 pcw developmental stages identified MERTK+ macrophage and DKK3+ and POSTN+ fibroblast cell populations that were predicted to correspond to equivalent populations identified in the adult tissues. In developmental tissues, MERTK + LYVE1 + MRC1+ macrophages localised to the shoulder capsule lining adjacent to GAS6+ fibroblasts as observed in adult tissues. The identification of MKI67 + TOP2A+ cycling myeloid cells supports the concept that myeloid cells divide and populate the soft tissues of the shoulder joint during foetal development, suggesting that MERTK+ macrophages are an embryonically seeded population. Adult frozen shoulder patient tissues also express Ki67, further supporting the concept of a self-sustaining macrophage population. The developmental origin of macrophages is well documented in visceral tissues including brain, heart, lung and liver48,49 and the pool of LYVE1 + FOLR2+ macrophages are maintained through self-renewal with minimal monocyte input50. However, knowledge of the distinct immune populations occupying the soft tissues of the joint during human embryonic development is limited. Our findings demonstrate a possible embryonic origin for the cell types implicated in matrix remodelling of the fibrotic niche, suggesting that a template to resolve adult fibrotic disease might be laid down during foetal development. As ontological processes are frequently recapitulated in inflammatory diseases, we hypothesize that frozen shoulder could exemplify a recapitulation of ontogeny and during disease the re-activation of developmental cellular programmes could drive fibrosis resolution.
동결견에서 MERTK+ 대식세포와 기질 관련 섬유아세포 간의 교신이 기질 재구성의 기반이 된다는 점을 확인한 후, 우리는 성인 섬유성 관절낭 질환의 해결에 관여하는 세포 유형이 태아 발달 기간에도 존재하는지 조사하였다. 12~17주 태아기 인간 어깨 조직의 단일 세포 분석을 통해, 성인 조직에서 확인된 동등한 집단과 일치할 것으로 예측되는 MERTK+ 대식세포 및 DKK3+ 및 POSTN+ 섬유아세포 집단이 확인되었다. 발달 중인 조직에서, MERTK+ LYVE1+ MRC1+ 대식세포는 성인 조직에서 관찰된 것과 마찬가지로 GAS6+ 섬유아세포에 인접한 어깨 관절낭 내벽에 국소화되어 있었다. MKI67+ TOP2A+ 분열 중인 골수계 세포의 확인은 태아 발달 기간 동안 골수계 세포가 분열하여 어깨 관절의 연조직에 정착한다는 개념을 뒷받침하며, MERTK+ 대식세포가 배아기 시드된 집단임을 시사한다. 성인 동결견 환자 조직에서도 Ki67 발현이 관찰되어 자가 유지되는 대식세포 집단의 개념을 추가로 지지한다. 대식세포의 발생학적 기원은 뇌, 심장, 폐, 간 등 내장 조직에서 잘 기록되어 있으며48,49 LYVE1+ FOLR2+ 대식세포 군집은 최소한의 단핵구 유입을 통한 자가 재생으로 유지됩니다50. 그러나 인간 배아 발달 과정에서 관절 연조직을 점유하는 특이적 면역 군집에 대한 지식은 제한적입니다. 본 연구 결과는 섬유화 미세환경의 기질 재형성에 관여하는 세포 유형의 배아 기원을 시사하며, 성인 섬유화 질환 해결의 틀이 태아 발달 기간에 마련될 수 있음을 제안한다. 발생학적 과정이 염증성 질환에서 빈번히 재현되는 점을 고려할 때, 동결견은 발생 과정의 재현을 예시할 수 있으며, 질환 진행 중 발달적 세포 프로그램의 재활성화가 섬유화 해결을 촉진할 수 있다고 가정한다.
Our study builds upon previous work highlighting the importance of distinct phases of inflammation, resolution and remodelling in diseases of musculoskeletal soft tissues51, generating new therapeutic targets that have the potential to exploit the cell-cell interactions we have observed in this uniquely self-limiting musculoskeletal condition. We have discovered a population of pro-inflammatory MERTKlowCD48+ macrophages that may be involved in disease pathogenesis and provide functional evidence in support of a role for MERTK+ macrophages in disease resolution. We identify MERTK, LYVE1, MRC1 and DKK3+ and POSTN+ fibroblasts as specific targets for functional studies of fibrosis in murine models. Therapeutic enhancement of the functions of MerTK+Mφ and augmenting integrin mediated cell-matrix interactions between MERTK + Mφ and DKK3+ and POSTN+ fibroblasts to regulate inflammation and induce matrix remodelling could accelerate resolution of frozen shoulder and resolve persistent inflammatory fibrotic pathologies affecting other tissues.
본 연구는 근골격계 연조직 질환에서 염증, 해결, 재구성의 각기 다른 단계의 중요성을 강조한 기존 연구를 기반으로 하며51, 이 독특한 자가제한성 근골격계 질환에서 관찰된 세포 간 상호작용을 활용할 잠재력을 지닌 새로운 치료 표적을 제시한다. 우리는 질병 발병 기전에 관여할 수 있는 염증성 MERTKlowCD48+ 대식세포 집단을 발견했으며, MERTK+ 대식세포가 질병 해결에 역할을 한다는 기능적 증거를 제시한다. 우리는 MERTK, LYVE1, MRC1 및 DKK3+와 POSTN+ 섬유아세포를 생쥐 모델에서 섬유화의 기능적 연구를 위한 특정 표적으로 확인하였습니다. MerTK+Mφ의 기능을 치료적으로 강화하고 MERTK+Mφ와 DKK3+ 및 POSTN+ 섬유아세포 사이의 인테그린 매개 세포-기질 상호작용을 증강시켜 염증을 조절하고 기질 재구성을 유도하면 동결견의 해결을 가속화하고 다른 조직에 영향을 미치는 지속적인 염증성 섬유화 병리를 해결할 수 있을 것입니다.
Methods
Study approval
The University of Oxford Research Governance & Ethics Assurance approved the protocol for this study. Ethical approval for the use of adult tissues for this study was granted by the local research ethics committee (ICECAP study REC reference 18/SC0649, IRAS project ID 257757) and the Oxford Musculoskeletal Biobank (19/SC/0134). Full written informed consent according to the Declaration of Helsinki was obtained from all patients. Foetal tissue samples were provided voluntarily with appropriate written informed consent, the Human Developmental Biology resource (HDBR) tissue bank operates under Research Ethics Committee approvals 18/NE/0290 (Newcastle upon Tyne) and 18/LO/0822 (London).
Statistics & reproducibility
The objective of this study was to utilise a naturally occurring human disease model of self-limiting fibrosis (advanced-stage frozen shoulder) to understand the cellular and molecular basis underpinning successful fibrosis resolution. Adult tissue samples used for this study were collected from the rotator interval of the shoulder capsule from well-phenotyped comparator and diseased patient cohorts. Patients with frozen shoulder were included in the study if they had a diagnosis of frozen shoulder ≥12 months. Patients with concurrent rheumatoid arthritis, osteoarthritis and those receiving dexamethasone treatment 3 months prior to surgery were excluded from the study. Foetal shoulder tissues were collected from between 12-17 post conception week developmental stages. We performed single-cell RNA sequencing to identify the cell types comprising the shoulder capsule during developmental, adult comparator and adult diseased states. Functional experiments using micro-cultures of adult frozen shoulder patient-derived cells were studied to identify the cell types, molecules, genes and biological pathways underpinning fibrosis resolution in vitro. Statistical analysis and sample size justification were derived from previous studies (24, 49) that were sufficiently powered to gain insights from studying inflammatory and resolving processes in human musculoskeletal tissues. For single-cell RNAseq and bulk RNAseq data, investigators were not blinded to allocation during experiments and outcome assessment. Quantitative analysis of histological capsule tissues sections was performed by a single investigator who was blinded to the health status of the shoulder capsule tissue. For immunohistochemistry, H&E staining was first performed on sequential tissues to determine the region of interest in imaging, a minimum of 5 regions from each tissue slide were analysed. Representative images are shown for micrographs in Figs. 1A, 2A, 2C, 3D, 3H, 6G–J and supplementary figures S3F–H, S5F, S6A–D, S7F–G, S9D & S10A–H. Representative images are shown from a minimum of 3 independent experiments in each instance. The details of sample size and statistical tests employed in each case were provided in the figure captions. All P values were corrected for multiple testing and the statistical testing method was indicated in the figure captions. We used the following convention to indicate significance with asterisks: not significant (ns) (P > 0.1), * (0.1 > P > 0.01), ** (0.01 > P > 0.001), ***(0.001 > P > 0.0001). No data were excluded from the analyses. The experiments were not randomized. Further information on research design is available in the Nature Reporting Summary linked to this article.
Collection of adult shoulder capsule tissues
Comparator and frozen shoulder adult tissue biopsies were collected from the rotator interval of the shoulder (glenohumeral) joint capsule. Comparator tissues were collected from male and female patients undergoing elective shoulder stabilisation (n = 8 donors) or shoulder arthroplasty surgical procedures (n = 12 donors). Frozen shoulder tissues were collected from patients with advanced-stage disease (≥12 months symptom duration) undergoing surgical arthroscopic capsular release (n = 15 donors) (Supplementary Data 1). As frozen shoulder is a condition that affects males and females similarly, we ensured adequate representation of both sexes in comparator and frozen shoulder patient cohorts. All experiments included tissues or cells derived from both male and female donors.
Processing of adult shoulder capsule tissues for scRNAseq
To identify the cell types comprising the resolving fibrotic niche, patient shoulder capsule tissue biopsies (n = 6 comparator, n = 4 frozen shoulder donors) were disaggregated and digested in DMEM F12 media containing 4 mg/mL Worthington Collagenase II (Lorne) and 1 mg/mL DNase (Lorne) at 37 °C for 90 mins with gentle agitation. The cell suspension was filtered and transferred to DMEM containing 10% foetal bovine serum (Gibco).
Processing of foetal shoulder joint tissues for scRNAseq
Upper limbs (n = 6) from male and female foetuses were collected by the Human Developmental Biology Resource (HDBR) team following medical or surgical termination of pregnancy (Supplementary Data 1). Developmental stage was determined by anthropometric parameters and number of somites present. Collected tissue samples were stored in L-15 medium at 4 °C during shipment, time between collection and subsequent processing was <5 h. Soft tissues of the shoulder joint were isolated, disaggregated and digested using Liberase TL (Roche) 0.1 mg/ml diluted in DMEM (Gibco) over 2 h at 37 °C. Liberated cells were collected every 20 minutes during digestion, followed by replacement of digestion media. The isolated cell suspension was filtered and transferred to DMEM containing 10% foetal bovine serum (Gibco).
scRNAseq of adult and developmental shoulder tissues
Single cell suspension samples were stained with 7-AAD dye (Bio-Legend 420404) for live/dead sorting on a SONY SH800 cell sorter. The density of cell suspensions was determined with the Bio-Rad TC20 Automated Cell Counter before loading 20,000 – 30,000 viable cells per sample onto a Next GEM Chip G and running on the 10x Chromium Controller. Single cell gene expression libraries were prepared using the 10x Chromium Next GEM Single Cell 3’ Reagent Kits v3.1 following manufacturer user guide (CG000204). The final gene expression libraries were sequenced on the Illumina NovaSeq6000 platform (v1.5 chemistry, 28 bp Read1 and >91 bp Read2) to a minimum depth of 50,000 reads per cell.
Processing and DAB immunostaining of adult shoulder capsule tissues
Shoulder capsule tissue biopsies collected from the rotator interval of comparator and frozen shoulder patients were immersed in 10% buffered formalin for a minimum of 48hrs. After fixation, samples were processed using a Leica ASP300S tissue processor and embedded in paraffin wax. Tissues were sectioned at 6μm using a rotary RM2135 microtome (Leica Microsystems Ltd) onto adhesive glass slides and baked at 60 °C for 30 mins and 37 °C for 60 mins. We used immunohistochemistry to validate key sub-population markers for macrophages and fibroblasts identified in Fig. 1. Comparator and frozen shoulder capsular tissue sections were obtained through deparaffinization and target retrieval steps (high pH, heat-mediated antigen retrieval) using an automated PT Link (Dako). Antibody staining was performed using the EnVision FLEX visualization system with an Autostainer Link 48 (Dako). Antibody binding was visualized using FLEX 3,3′-diaminobenzidine (DAB) substrate working solution and hematoxylin counterstain (Dako). Details of antibodies and their working dilutions are shown in Supplementary Table 1. For negative controls, the primary antibody was substituted for universal isotype control antibodies: cocktail of mouse immunoglobulin G (IgG1), IgG2a, IgG2b, IgG3, and IgM (Dako) and rabbit immunoglobulin fraction of serum from nonimmunized rabbits, solid-phase absorbed (Dako). Isotype control images are shown in Figure S6A-D. After staining, slides were taken through graded industrial methylated spirit and xylene, mounted in DPX mounting medium (Fischer Scientific) and imaged on a Brightfield microscope (Olympus). Images were acquired on a MOTIC slide scanner (Leica Biosystems) by a single blinded investigator. Image analysis was conducted using FIJI (National Institutes of Health version 2.3.0) using previously validated methodology52. For each sample, immunopositive staining was normalized to the number of hematoxylin-counterstained nuclei within the field of view. Data are presented as % area immunostaining. Statistical analyses were performed with GraphPad Prism, version 9.2.0 (GraphPad Software). Statistically significant differences in immunopositive staining were calculated using the Pairwise Mann Whitney U test. P < 0.05 was considered statistically significant.
Validation & spatial mapping of distinct macrophage & fibroblast subsets in adult capsular tissues
We performed multiplex immunostaining to validate the phenotype of identified macrophage and fibroblast clusters and to identify the topographical niches these cells occupy in comparator and frozen shoulder adult capsular tissues using a previously validated protocol51. After antigen retrieval steps, tissues were blocked in 5% normal goat serum (Sigma) in phosphate-buffered saline (PBS) for 30 mins in a humid chamber at room temperature. Sections were incubated with the primary antibody cocktail diluted in 5% normal goat serum in PBS for 2.5 hrs at room temperature (primary antibodies used are listed in Supplementary Table 1A). Sections were washed with PBS–Tween 20 (PBST) and incubated in the secondary antibody cocktail, each diluted 1:200 in 5% normal equine serum (Sigma) in PBS for 2.5 hrs. The secondary antibodies were Alexa Fluor goat anti-mouse IgG2a or IgG2b or goat anti-rabbit IgG (Life Technologies) and goat anti-mouse IgG1 (Southern Biotech) (Supplementary Table 1B). After washing, sections were incubated in 2 mM POPO-1 nuclear counterstain (Life Technologies) diluted in PBS containing 0.05% saponin (Sigma) for 20 mins. Tissue autofluorescence was quenched with a solution of 0.1% Sudan Black B (Applichem) in 70% ethanol for 3 mins. Slides were mounted using fluorescent mounting medium (VectaShield), sealed, and stored at 4 °C until image acquisition. For negative controls, the primary antibody was substituted for universal isotype control antibodies: cocktail of mouse IgG1, IgG2a, IgG2b, IgG3, and IgM (Dako) and rabbit immunoglobulin fraction of serum from non-immunized rabbits, solid-phase absorbed (Dako) (Supplementary Table 1B). Isotype control images are shown in Fig S6C & D. Images were acquired on a Zeiss LSM 710 confocal microscope using a 40x oil immersion objective (numerical aperture, 0.95). The fluorophores POPO-1, FITC, Alexa Fluor 568, and Alexa Fluor 633 were excited using the 405, 488, 561, and 633 nm laser lines, respectively. To minimize bleed-through, all channels were acquired sequentially. Averaging was set to 2 and the pinhole was set to about 1 airy unit. Two-dimensional image reconstructions were created using ZEN Black software (Zeiss version 11.0.3.190).
Validation of T cell subsets in frozen shoulder patient tissues
Multiplex ChipCytometry staining was used for immunophenotyping of T cell populations in frozen shoulder patient tissues using a previously validated protocol53. Cryosections were cut directly onto APES-coated coverslips (Sigma-Aldrich) and fixed immediately using freshly prepared 0.1 M phosphate-buffered 4% paraformaldehyde (Sigma-Aldrich) or Zellkraftwerk fixation buffer (Zellkraftwerk) for 10 mins at room temperature. After washing in PBS, sections on coverslips were assembled into tissue chips (ZellSafe Tissue – Chips; Zellkraftwerk). Tissue sections were blocked by incubating in 5% normal goat serum (Thermo Fischer Scientific) in PBS for 1 hr at room temperature. Immunostaining was performed at room temperature for 30 mins using 0.5 ml of Ab solution per chip. Antibody cocktails were diluted in PBS alone or PBS containing 2% normal goat serum. Markers were acquired in iterative rounds of photobleaching, staining, and imaging including CD45, CD3, CD4, CD8, GZMB, GZMK, CD127, CD40LG, CD18, CD161, CD2, CD5, FAP, CD31 (Supplementary Table 2). Images were downloaded from the proprietary software (Zellscanner App, version 19.08.2020, Zellkraftwerk Gmbh) as single marker 16-bit greyscale tif files. An outlier filter was applied to reduce signal noise (median, radius 0.5) and images were cropped, merged and colourised in FIJI (version 2.3.0) using previously validated methodology54.
Processing and immunostaining of developmental shoulder joint tissues
Foetal shoulders were fixed in 10% formalin for a minimum of 48hrs. Samples >10 post conceptional weeks were decalcified with 0.5 M EDTA solution for 15-30 days (depending upon developmental stage). Subsequently, samples were processed into paraffin using a tissue processor (Tissue TeK VIP 6 Processor, Sakura) and embedded in paraffin blocks in anteroposterior orientation. Tissues were sectioned at 6μm as per adult tissues. Multiplex immunostaining was performed to validate the phenotype and topographical niches of the major cell types comprising the shoulder joint capsule during development. Sections of foetal shoulder tissues were baked overnight at 60 °C. Slides were transferred to xylene for de-waxing followed by a series of rehydration steps in ethanol solutions (100%, 95%, 70%, 50%). Each step was repeated twice for 5 mins each, followed by 2 washing steps in 1x PBS. The slides were then permeabilised for 10 mins in 0.3% Triton X-100 and washed further in 1x PBS for 5 mins. Heat-induced Epitope Retrieval (HIER) was conducted with the NxGen decloaking chamber (Biocare Medical) with both Citrate (pH 6.0) and Tris (pH 9.0) antigen retrieval solutions. The decloaking chamber is programmed to incubate slides in Citrate solution (Vector Labs, H3300) for 20 mins reaching 110 °C and 6.1 PSI, which was maintained for 4 mins before cool down was initiated. Slides were transferred to a Tris solution for 20 mins followed by an additional 10 mins at room temperature (RT). Tissue slides were blocked with a 3% BSA (Merck, A7906) and 10% Donkey serum (Bio-Rad, C06SB) solution overnight at 4 °C. Slides were washed in 1xPBS for 10 mins and then stained with DAPI (Thermo, D3571) for 15 mins. Slides were washed in 1xPBS for 5 mins and coverslipped with 75 µl of mounting media (50% glycerol – Sigma, G5516 and 4% propyl gallate – Sigma, 2370).
Cell DIVE Imaging of adult & developmental shoulder capsule tissues
The GE Cell DIVE system was used to image adult and developmental FFPE slides using the ImageApp imaging software (version 1.0)55,56. An initial image using a 10x objective of the entire tissue allows for region of interest (ROI) selection. The background and innate autofluorescence of the tissue are captured during the initial 20x imaging round. This uses the FITC, Cy3, Cy5 channels as well as the DAPI signal to also create a virtual H&E image. Background imaging is used to subtract autofluorescence from all subsequent rounds of staining. Slides were de-coverslipped in 1xPBS prior to staining. Each staining round consisted of a master mix of 3 antibodies prepared in antibody diluent (3% BSA, 1xPBS). The initial round used primary antibodies which were incubated in the dark for 1 hr at room temperature followed by 3x washes in 1xPBS. Secondary antibodies raised in Donkey were then incubated for an additional hour at room temperature which were either conjugated to Alexa Fluor 488, 555 or 647. Each subsequent staining round used directly conjugated antibodies to either of these dyes. Fluorophores were bleached between each staining round using NaHCO3 (0.1 M, pH 11.2. Sigma - S6297) and 3% H2O2 (Merck – 216763). Fresh bleaching solutions were prepared, and slides were bleached 3 times (15 minutes each) with a 1 minute 1xPBS wash in between bleaching rounds. Slides were re-stained for DAPI for 2 minutes and washed in 1xPBS for 5 minutes before imaging the dye-inactivated round as the new background round (for subsequent background subtraction). DAPI staining between imaging rounds assists in image registration and alignment. Slides were multiplexed with the next panel of three markers with iterative staining, bleaching and imaging. A negative control slide stained with isotype-matched antibodies was used at the same concentration as the corresponding primary antibodies and using the same exposure settings. Antibodies used for Cell DIVE immunostaining are listed in Supplementary Table 3. QuPath software (version 0.3) was used for image visualisation and the isotype controls were contrast-matched to samples stained with primary antibodies for comparison57.
Quantitative analysis of Cell DIVE staining in adult shoulder capsule tissues
To determine the relative proportions of cell types between comparator and frozen shoulder patient tissues, we performed Cell DIVE multiplex immunofluorescence staining for CD31, CD68 and DKK3 on comparator (n = 5 donors) and frozen shoulder (n = 5 donors) tissue sections. These markers were used to identify vascular endothelial cells, macrophages and fibroblasts, respectively. To quantify the result, we selected regions of interest from the staining and developed an analysis pipeline with custom code written in Matlab (version R2023a) and CellProfiler (version 4.2.5) to divide ROIs into tiles about cell nuclei (averaging 1058 tiles per ROI), then assign cell types to individual tiles from staining intensity. In brief, this pipeline tessellated ROIs using the Voronoi algorithm and DAPI nuclear staining to subdivide ROIs into multiple regions or “tiles”. Each tile contains a single nucleus, and the tile borders are equidistant between neighbouring nuclei as a result of the Voronoi algorithm. The normalized highest intensity signal in each tile was calculated to assign each tile a state of either CD31+, CD68+ or DKK3 + , and the results shown in Figure S1E. This approach was used to minimise uncertainty from differences in cell size.
Isolation of monocyte-derived macrophages from blood
Non-clinical blood cones were provided by the NHSBT and approved under REC number 11/H0771/7. PBMC were isolated using a density gradient (Histopaque, Sigma) and depleted from red blood cells using ACK lysis buffer (ThermoFisher Scientific). CD14+ CD16+ cells were isolated from peripheral blood mononuclear cells using EasySep™ Human Monocyte Enrichment Kit without CD16 Depletion (Stemcell) according to the manufacturer’s protocol. Cells were differentiated into macrophages in complete media (RPMI, Gibco + FCS, Gibco +1%Penstrep, Sigma) containing 5% FCS and additional 100ng/mL M-CSF (PeproTech) with incubator conditions 37 °C/5% CO2. On day 6, macrophages were harvested using cell lifters and plated in the appropriate plate at a density of 500,000 cells/well in a 6-well plate or 100,000 cells/well in a 96-well plate, in 3 mL and 200uL complete media, containing 3% FCS respectively and rested for 2 days. On day 8, cells were washed and stimulated with 10 ng/mL LPS (Invivogen) or 1uM Dexamethasone (Sigma) for 72 hrs. Doses of reagents used were adapted from a previously published protocol24.
Characterisation of Dexamethasone and LPS treated MDMs by flow cytometry
To characterise the blood cone derived macrophages after stimulating with Dexamethasone or LPS, protein expression was measured by flow cytometric analysis. For this experiment 4 different donors were used in independent experiments. Monocytes were isolated from blood cones and treated with M-CSF for 5 days as described above. After plating to 1×106, cells were rested for 2 days and stimulated with LPS or Dexamethasone to induce respective MerTKlow or MerTKhigh phenotypes as above. Macrophages were incubated with Accutase (Invitrogen) for 10 mins then harvested with a cell lifter for flow cytometry. The cells were stained with antibodies against the surface markers MerTK, CD163, CD206, TREM2, CD14, LYVE-1 and CD48 (1:100 dilution, Supplementary Table 4A). In addition, a Live/dead stain (1:500) and FC block (1:100) was added to the mixture. After 20 mins of staining at 4 degrees, cells were centrifuged, and supernatant was discarded before cells were fixed using 4% PFA in PBS at RT for 20 mins. After centrifugation and discarding of the supernatant, cells were permeabilized for 15 mins with 1x BD Perm/Wash™ buffer and stained with antibody against CD68 for 20 mins at RT. Cells were washed in permeabilization buffer and resuspended in FACS buffer (10% FBS in PBS) before running on an LSRII Flow Cytometer. Data was analysed using FlowJo V10. Data were expressed as geometric mean of staining intensity and were normalised within donors relative to control (minus Dexamethasone or LPS stimulation). Data (from n = 4 donors) were analysed in GraphPad Prism (version 9.2.0). Data are expressed as SEM, statistical significance was determined using an unpaired t-test, P < 0.05. Relative protein expression of MerTK, CD206 and CD48 across 4 donors is shown in Figure S7C, representative contour plots showing expression of MerTK, CD206 and CD163 are shown in Figure S7D.
Coculture of MerTKhigh and MerTKlow MDMs with primary capsular fibroblasts
Monocyte derived macrophages (MDMs, n = 3 donors) in 6-well plates were stimulated for 72 hrs with the previously described conditions to induce MerTKhigh or MerTKlow populations. After washing with PBS, capsule-derived fibroblasts isolated from frozen shoulder patients (n = 3 donors) were added to MerTKhigh or MerTKlow incubations at a density of 200,000 cells/well. Fibroblasts and MDMs were co-cultured for 48 hrs in complete media containing 3% FCS prior to harvest and sorting (Figure S7E). After 48 hrs, cells were incubated with Accutase for 10 mins and vigorously resuspended. Media was added and remaining adherent cells detached by scraping. A single cell suspension was obtained after putting cells through a 70 μm cell strainer and resuspended in Facs buffer containing 1% BSA 0.1 mg/ml DNase in PBS. Cells were stained with antibodies PDPN, CD90, CD45, CD14 (1:200 dilution, Supplementary Table S4B) to distinguish between fibroblasts and MDMs. Directly co-cultured MDMs and fibroblasts were sorted on a BD Aria III with Diva 8.01 software. Fibroblasts (PDPN + CD90 + CD31-) and MDMs (CD45 + CD14 + ) were sorted into RLT Lysis buffer (Qiagen). RNA from sorted capsular fibroblasts was extracted using the RNEasy microprep kit (Qiagen) according to the manufacturers protocol.
Bulk RNA-seq of capsular fibroblasts co-cultured with MDMs
Total RNA was quantified using Quant-it RiboGreen RNA Assay Kit (Invitrogen), and the integrity assessed by the 4200 Tapestation system (Agilent). PolyA+ mRNA was enriched and purified from 100 ng high-quality total RNA (RIN > 9) using NEBNext Poly(A) mRNA Magnetic Isolation Module (E7490L). Generation of double stranded cDNA and library construction were performed using the NEBNext Ultra II Directional RNA Library Prep Kit for Illumina (E7760L), with custom adapters and barcode tags (dual indexing, based on https://doi.org/10.1186/1472-6750-13-104). Indexed libraries were multiplexed based on fluorescent-based quantification. The final size distribution of the multiplexed pool was determined using Tapestation and quantified by Qubit assay (Thermo Fisher), before sequencing on an Illumina NovaSeq6000 v1.5 in 150 bp paired-end mode. Data from the NovaSeq6000 were demultiplexed using bcl2fastq (Illumina) version 2.20.0. Cassette scores were calculated using addModuleScore function and the log normalised count of all MERTKhigh markers.
Immunocytochemistry for markers of matrix-associated fibroblasts
Human monocyte-derived macrophages were seeded in chamber slides and stimulated for 72 hrs to drive either a MerTKlow or MerTKhigh phenotype as described above, along with unstimulated ‘M0’ monocyte-derived macrophages as control. Following stimulation, primary frozen shoulder fibroblasts (n = 3 donors) were co-cultured with macrophages for 48 h prior to fixation with 4% PFA. Cells were permeabilized with 0.1% Triton-X, blocked with 5% normal goat serum in PBS + 0.1% Tween-20 (PBS-T), then stained with POSTN (1:200, abcam #ab79946) and DKK3 (1:400, Proteintech #66758) overnight, followed by secondary antibodies at 1:200 (goat anti-mouse 633, goat anti-rabbit 568, Life Technologies). Nuclei were counterstained with POPO-1 (1:1000, ThermoFisher #P3580) in 0.05% saponin, and slides imaged using an inverted Zeiss LSM 710 confocal microscope with 10X objective. Respective isotype control staining is shown in Figure S7G.
Computational analysis of single-cell RNA-sequencing dataAnalysis of single-cell RNA-seq data from frozen shoulder and comparator adult tissues
Single cell RNA sequencing data was aligned to human genome using 10x Genomics Cell Ranger version 6.1. and 2020-A Reference annotations (GRCh38, GENCODE v32/Ensembl 98). Ambient background RNA was removed using CellBender (version 0.2.0; droplets included=10,000, fpr=0.01, epochs=150). A total of n = 13194 cells were identified for downstream analysis. Doublets were determined using Scrublet (version 0.2.3) at an expected doublet rate of 0.0558. The count matrix was transformed into a Seurat object for downstream analysis59. Additionally, we removed cells that had abnormally low or high gene counts ( < 300 or >6500 n_Feature) and high mitochondrial gene expression ( > 10% mitochondrial genes, n = 5818 cells removed). Cell cycle scores were computed using curated gene sets60 and the addModuleScore function provided in Seurat (version 4.0.4). Cell counts were log normalised, scaled, and n = 3000 highly variable genes (HVG) were selected using the VST method and used for principal component analysis (PCA, n = 50 PCs retained). We corrected the latent space using Harmony by the condition (i.e., comparator and frozen shoulder) of the samples (n = 15 components retained)15. Downstream analyses were performed using the Cellhub workflow (https://github.com/sansomlab/cellhub). Nearest neighbour graphs were computed with the HNSW algorithm (Euclidean distance, k = 20) and clustering performed with the Leiden algorithm. We applied additional filters to remove clusters of cells with low UMI (n_Count <10000) and gene count (n_Features <1000), and highly expressed hemoglobin markers (HBA, HBB, HBG) (n = 558 cells). The remaining clusters of high-quality cells (n = 6818 in total) were assigned to stromal, myeloid or lymphoid cell type “regions” based on singleR cell type predictions61 (version 1.06) and expression of known cell-type specific marker genes (CD34 PECAM–, stromal; CD14, myeloid cells; CD3, lymphoid cells). The count matrices for each of the cell-type regions were extracted and separately re-analysed using the workflow described above. For the stromal cells we used n = 1500 HVG, n = 50 PCA components, n = 14 harmony components and a clustering resolution of 0.3. For the myeloid cells we used n = 400 HVG, n = 50 PCA components, n = 11 harmony components and a clustering resolution of 0.2. We removed a cluster of cells that consisted mainly doublets (score ≥ 0.4; n = 9 cells removed). For the lymphoid cells n = 1000 HVG, n = 50 PCA components, n = 11 harmony components and a clustering resolution of 0.2 was used. The markers of each cluster were identified using a Wilcox test (BH adjusted P ≤ 0.05).
Pseudobulk-level differential expression analysis
Within cell type differential expression analyses of the adult single cell dataset (comparator vs frozen shoulder) were performed at pseudo-bulk level. Pseudo-bulks were created by summing gene counts within cluster for each sample using Muscat. Differential expression analysis was performed using DESeq262 (version 1.26.0; ashr shrinkage, Wald test). Genes expressed (≥10 counts) in n ≥x samples where x was equal to the number of biological replicates (Supplementary Data 3 & 4) in any of the condition groups were retained for differential expression analysis. To identify genes that were regulated differently in frozen shoulder and RA relative to healthy controls we performed an interaction analysis. To do so, we created pseudobulk datasets as described above and employed an interaction term (condition:tissue) in the regression model (~condition + tissue). The log2 Fold changes were adjusted using ashr shrinkage, and P-values were corrected for multiple testing (BH adjusted P ≤ 0.05). We used Wilcoxon test to compare the median log2 fold changes between tissue types, and calculated correlation coefficient using Spearman’s correlation.
Pathway analysis
To further dissect the functionality of genes and gene sets that had been identified from the sequencing data, we performed pathway over-representation analysis using Gene Ontology database63,64 and Canonical pathway databases (BioCarta, http://cgap.nci.nih.gov/Pathways/BioCarta_Pathways; KEGG, http://www.pathway.jp; Pathway Interaction Database, PID, http://pid.nci.nih.gov; Reactome, http://www.reactome.org and WikiPathways, https://www.wikipathways.org/) with gsFisher (https://github.com/sansomlab/gsfisher) and ClusterProfiler (version 4.2.2)65. For the analysis of the single cell sequencing data, we used marker genes (Wilcox, BH adjusted P ≤ 0.1) representative of each cluster and restricted the gene universe to genes that were detected in a minimum of 10% of cells. For the bulk sequencing data, we used the differentially expressed genes (DESeq2, BH adjusted P ≤ 0.05) and restricted the gene universe to include genes that were in the DEG analysis. Significant pathways were ranked and selected by their magnitude of the odds ratio, which was evaluated using Fisher Exact test and corrected for multiple testing using the Benjamini-Hochberg algorithm.
Cell-cell interaction analysis
We identified receptor-ligand interactions between cell clusters using Network Analysis Toolkit for Multicellular Interactions (NATMI)23, we retained only fully validated interactions from literature and the constituent ligand and receptor genes that are expressed at a detectable level (>5% within cluster).
Protein-protein and gene regulatory network analyses
Protein association network analysis was performed using Intact25 (release 242) and the resulting network graph was filtered to retain nodes with a maximum of two degrees of separation. Single-cell gene regulatory network analysis of the fibroblasts was performed using pySCENIC66 (version 0.11.2) and the elicited biomolecular interaction networks and regulons were visualized using iRegulon67 and Cytoscape68 (version 3.9.1) We used logistic regression to model the cassette score distribution in macrophage clusters between adult shoulder capsule and RA synovial tissues, which was statistically evaluated using Wald test.
Comparison of macrophages from frozen shoulder and knee synovium
Data (Seurat Object) for macrophages from the knee synovium was a gift from the corresponding authors of #PMID:3260133524 and can be retrieved from ArrayExpress under the accession E-MTAB-8322. We retained 19078 genes that had matching Ensembl ID in both datasets. We projected the annotations of the reference data, i.e., the adult shoulder capsule on to the cells from the knee synovium through semi-supervised machine learning using scANVI of scArches26 with condition as the batch variable. The reference data and query data were both trained for 400 epochs. We accepted the predicted label of each query cell based on the highest score for the cluster (we observed minimal differences when a threshold of 0.6 or 0.8 was applied, data not shown).
Analysis of single-cell RNA-seq data from foetal tissues
Single cell RNA sequencing data from foetal tissues were analysed using the same workflow as described in the adult atlas analysis section except that use of an integration algorithm was found not to be necessary. The foetal data comprised of n = 17718 cells, of which 4538 cells were removed for abnormal gene count and high mitochondria gene expression (threshold as before). We selected n = 6000 HVG and utilised the top n = 16 PCs for computation of the UMAP and nearest neighbour graph. After removing a further n = 519 cells that had a low gene count), low UMI count (threshold as before), HBB+ cells, we retained 12,661 high-quality cells. The count matrices for each of the cell-type regions were extracted and separately re-analysed. For the stromal cells we used n = 2000 HVG, n = 14 PCs and a clustering resolution of 0.7. For the myeloid cells we used n = 500 HVG, n = 11 PCs and a clustering resolution of 0.6. For T cells n = 1000 HVG, n = 7 PCs and a clustering resolution of 0.2 was used. For B cells n = 500 HVG, n = 7 PCs and a clustering resolution of 0.3 was used. The markers of each cluster were identified using a Wilcox test (BH adjusted P-value ≤ 0.05).
Computational analysis of bulk RNA-sequencing dataData processing and differential expression analysis
We assessed the quality of the paired-end bulk RNA sequencing data using fastQC and generated count matrix using release 32 of the Gencode transcripts reference panel (Grch38.p13) with Salmon69(version 1.5.2). We performed dimension reduction (plotPCA; DESeq2) to determine the validity of the sample labels, and the sources of variation, and to visualize the structure of the data after the batch effect (stimulation) was adjusted using Limma70 (removeBatchEffect version 3.50.0). Only genes that were expressed (counts≥10) in all the samples in any of the condition groups were kept for analysis (n = 17544 genes retained). Differential expression analysis was performed using DESeq262 (version 1.34.0 apeglm shrinkage), and the statistical significance between the full model (~sample + stimulation) and the reduced model (~sample) was evaluated using LRT.
Cell type deconvolution analysis
Deconvolution of the bulk RNA sequencing data to the adult single cell sequencing data was performed using MuSiC71 (version 0.2.0). We used expression dataset objects from the raw count matrix of the single cell data using BisqueRNA72 (version 1.0.5) and from the Bulk RNA sequencing data using Biobase73(version 2.54.0). Differences in proportion of subtypes of cultured fibroblasts were evaluated for statistical significance using Kruskal-Wallis, followed by pairwise post-hoc test with Dunn’s.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The single cell-RNAseq data generated in this study have been deposited in the ENA database under accession code ERP143358 (https://www.ebi.ac.uk/ena/browser/view/PRJEB58305). The bulk-RNAseq data generated in this study have been deposited in the ENA database under accession code ERP143359. Human reference genome GRCh38 (GENCODE v32 / Ensembl 98) was used in the alignment of the bulk and single cell transcriptomic data. Source data are provided with this paper.
Code availability
The code used to perform quantitative analysis of Cell DIVE images for Figure S1E is available at Zenodo https://zenodo.org/record/8138743. A lightweight interactive visualisation tool for the adult shoulder capsule scRNAseq data is available on https://capsule.ndorms.ox.ac.uk.
References