
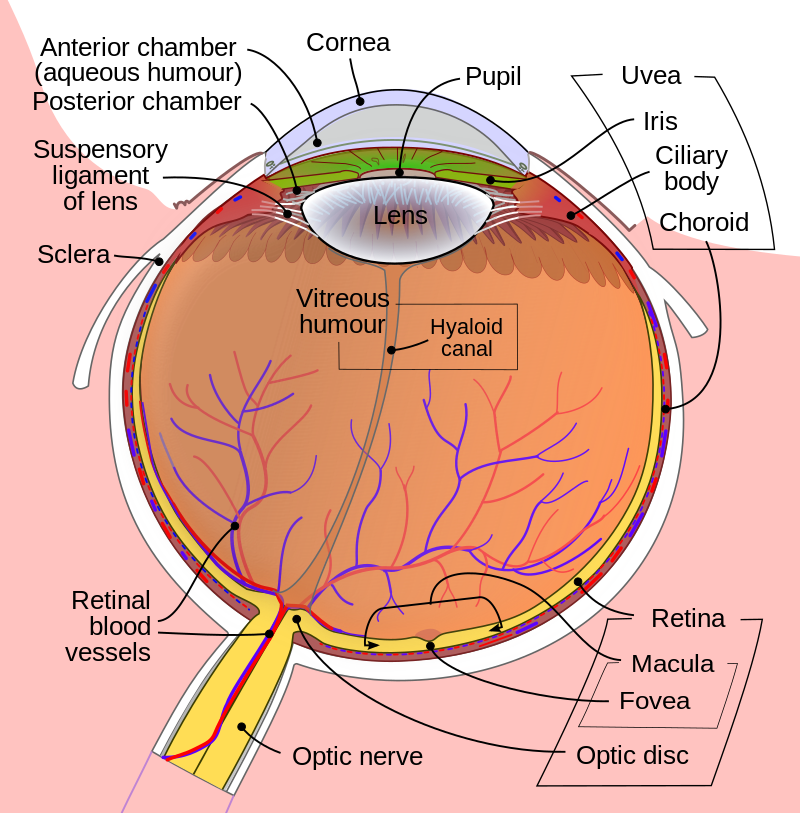
눈구조 해부학
Cornea 각막
uvea 포도막 - iris홍채, ciliary body 섬모체 choroid 맥락막
Sclera 공막
aqueous humor 방수 - anterior chamber 안구 앞방, posterior chamber 안구 뒷방
pupil 동공
lens 수정체 - suspensory ligament of lens 수정체 지지인대
vitreous humor - 유리체액(초자체액)
hyaloid canal - 유리체관
retina 망막 - macula 황반, fovea 망막중심오, optic disc 시신경 유두
retina ganglion cell 망막 신경절 세포
optic nerve 시신경
Cells. 2023 Dec; 12(24): 2797.
Published online 2023 Dec 8. doi: 10.3390/cells12242797
PMCID: PMC10741833
PMID: 38132117
The Role of Retinal Ganglion Cell Structure and Function in Glaucoma
Kathy Ming Feng,† Ta-Hsin Tsung,† Yi-Hao Chen, and Da-Wen Lu*
Paola Bagnoli, Academic Editor
Author information Article notes Copyright and License information PMC Disclaimer
Associated DataData Availability Statement
Abstract
Glaucoma, a leading cause of irreversible blindness globally, primarily affects retinal ganglion cells (RGCs). This review dives into the anatomy of RGC subtypes, covering the different underlying theoretical mechanisms that lead to RGC susceptibility in glaucoma, including mechanical, vascular, excitotoxicity, and neurotrophic factor deficiency, as well as oxidative stress and inflammation. Furthermore, we examined numerous imaging methods and functional assessments to gain insight into RGC health. Finally, we investigated the current possible neuroprotective targets for RGCs that could help with future glaucoma research and management.
전 세계적으로 돌이킬 수 없는 실명의 주요 원인인 녹내장은 주로 망막 신경절 세포(RGC)에 영향을 미칩니다.
이 리뷰에서는
기계적,
혈관적,
흥분 독성,
신경 영양 인자 결핍,
산화 스트레스 및 염증 등
녹내장에서 RGC 감수성을 유발하는 다양한 근본적인 이론적 메커니즘을 다루면서 RGC 하위 유형의 해부학을 자세히 살펴봅니다.
또한, 다양한 영상 촬영 방법과 기능 평가를 통해 RGC 건강에 대한 인사이트를 얻었습니다. 마지막으로, 향후 녹내장 연구 및 관리에 도움이 될 수 있는 RGC의 현재 가능한 신경 보호 표적에 대해 조사했습니다.
Keywords: retinal ganglion cells, glaucoma, neuroprotection, neurodegeneration, optical coherence tomography, imaging
1. Introduction
Glaucoma is a multifactorial eye disease defined by the progressive degeneration of retinal ganglion cells (RGCs) and their axons, eventually leading to irreversible vision loss. It has been projected that the worldwide prevalence of individuals affected by glaucoma will experience a substantial growth of 74% between the years 2013 and 2040 [1]. Glaucoma poses significant clinical and public health challenges as one of the leading causes of blindness around the globe. While elevated intraocular pressure (IOP) remains a significant risk factor and therapeutic target for glaucoma, it is becoming increasingly apparent that other factors may play a role in the disease’s pathogenesis and progression. The structure and function of RGCs, which serve as the ultimate output neurons of the retina and transmit visual information to the brain, are among the most crucial components being studied.
녹내장은
망막 신경절 세포(RGC)와 그 축삭의 점진적인 퇴행으로 정의되는 다인성 안과 질환으로,
결국 돌이킬 수 없는 시력 상실로 이어집니다.
2013년에서 2040년 사이에 녹내장 환자의 전 세계 유병률이 74%까지 크게 증가할 것으로 예상됩니다[1]. 녹내장은 전 세계 실명의 주요 원인 중 하나로서 임상 및 공중 보건에 심각한 문제를 야기하고 있습니다.
안압 상승이 녹내장의 중요한 위험 인자이자 치료 목표이지만, 녹내장의 발병과 진행에 다른 요인이 작용할 수 있다는 사실이 점점 더 분명해지고 있습니다. 망막의 최종 출력 뉴런 역할을 하며 시각 정보를 뇌로 전달하는 RGC의 구조와 기능은 가장 중요하게 연구되고 있는 요소 중 하나입니다.
The study of RGCs provides distinct insights not only into the mechanistic foundations of glaucoma but also into potential avenues for intervention and treatment. RGC apoptosis and axonal loss within the inner retina may implicate the earliest manifestation of glaucoma and exhibits a direct correlation with the clinical severity of the disease [2,3]. Apoptosis plays an essential role in maintaining homeostasis during normal development and aging; however, pathologic apoptosis is associated with age-related macular degeneration, retinitis pigmentosa, and neurodegeneration [4,5]. RGC damage can also be ascribed to various contributing mechanisms, including elevated IOP, impaired mitochondrial activity, oxidative stress, activation of glial cells, and excitotoxicity. With advances in imaging technology and functional assessment instruments such as optical coherence tomography, confocal scanning laser ophthalmoscopy, and adaptive optics, it is now possible to evaluate RGCs in greater detail [2]. A comprehensive understanding of how RGCs are affected by glaucoma could potentially transform the disease’s diagnosis and management, allowing ophthalmologists to provide more precise and personalized care to patients.
This review examines the structure and function of RGCs in the context of glaucoma. This includes topics ranging from the anatomy and physiology of RGCs to their susceptibility to glaucomatous conditions. In addition, this review investigates the most recent diagnostic imaging techniques and functional assessment methods. The review concludes by examining emerging therapeutic approaches that target RGCs, focusing on their clinical implications and future potential.
RGC에 대한 연구는
녹내장의 기계적 기초뿐만 아니라
개입 및 치료를 위한 잠재적인 방법에 대한 뚜렷한 통찰력을 제공합니다.
망막 내 RGC 세포 사멸과 축삭 손실은
녹내장의 초기 증상을 암시할 수 있으며
질병의 임상적 중증도와 직접적인 상관관계를 보입니다 [2,3].
세포 사멸은
정상적인 발달과 노화 과정에서 항상성을 유지하는 데 필수적인 역할을 하지만,
병적 세포 사멸은
연령 관련 황반변성,
망막색소변성 및
신경 퇴행과 관련이 있습니다 [4,5].
age-related macular degeneration, retinitis pigmentosa, and neurodegeneration
RGC 손상은
안압 상승,
미토콘드리아 활동 장애,
산화 스트레스,
신경교세포 활성화,
흥분성 독성 등 다양한 기전으로 인해 발생할 수 있습니다.
elevated IOP, impaired mitochondrial activity, oxidative stress, activation of glial cells, and excitotoxicity.
빛간섭단층촬영, 공초점 주사 레이저 검안경, 적응광학 등 영상 기술과 기능 평가 기기의 발전으로 이제 RGC를 더욱 자세히 평가할 수 있게 되었습니다[2]. RGC가 녹내장에 어떻게 영향을 받는지에 대한 포괄적인 이해는 안과 전문의가 환자에게 보다 정밀하고 개인화된 치료를 제공할 수 있도록 녹내장의 진단과 관리에 잠재적으로 변화를 가져올 수 있습니다.
이 리뷰에서는 녹내장의 맥락에서 RGC의 구조와 기능을 살펴봅니다. 여기에는 RGC의 해부학 및 생리학부터 녹내장 질환에 대한 감수성까지 다양한 주제가 포함됩니다. 또한 이 리뷰에서는 가장 최근의 진단 영상 기술과 기능 평가 방법을 조사합니다. 이 리뷰는 RGC를 표적으로 하는 새로운 치료 접근법을 임상적 의미와 향후 잠재력에 초점을 맞추어 검토하는 것으로 마무리합니다.
2. Basic Anatomy and Function of Retinal Ganglion Cells
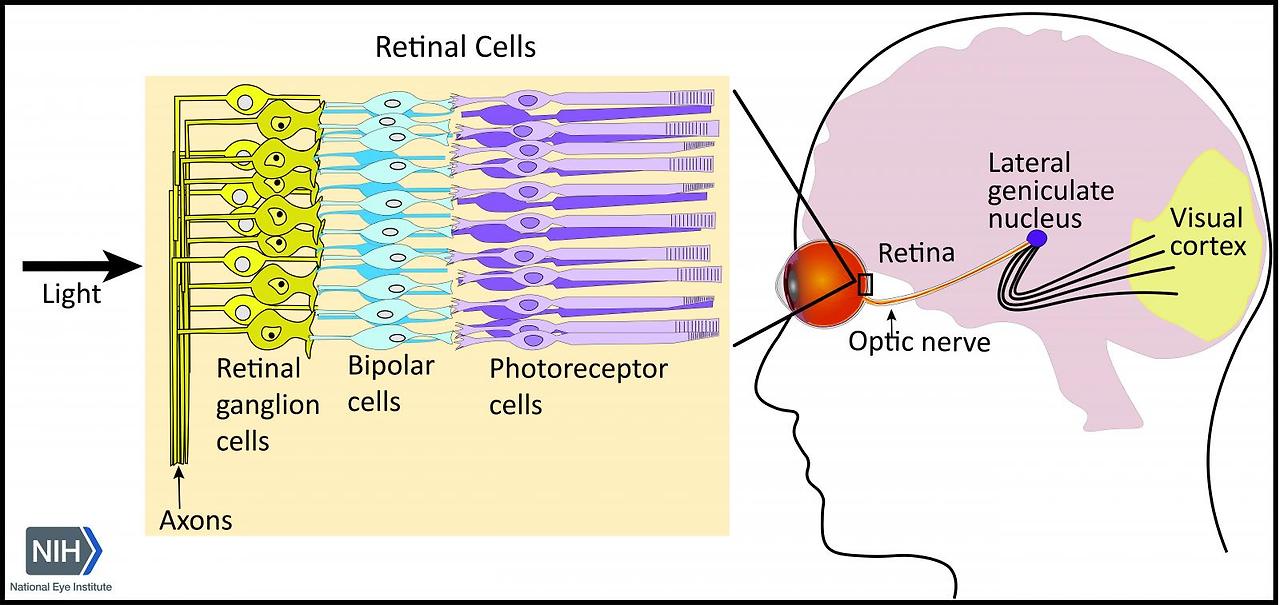
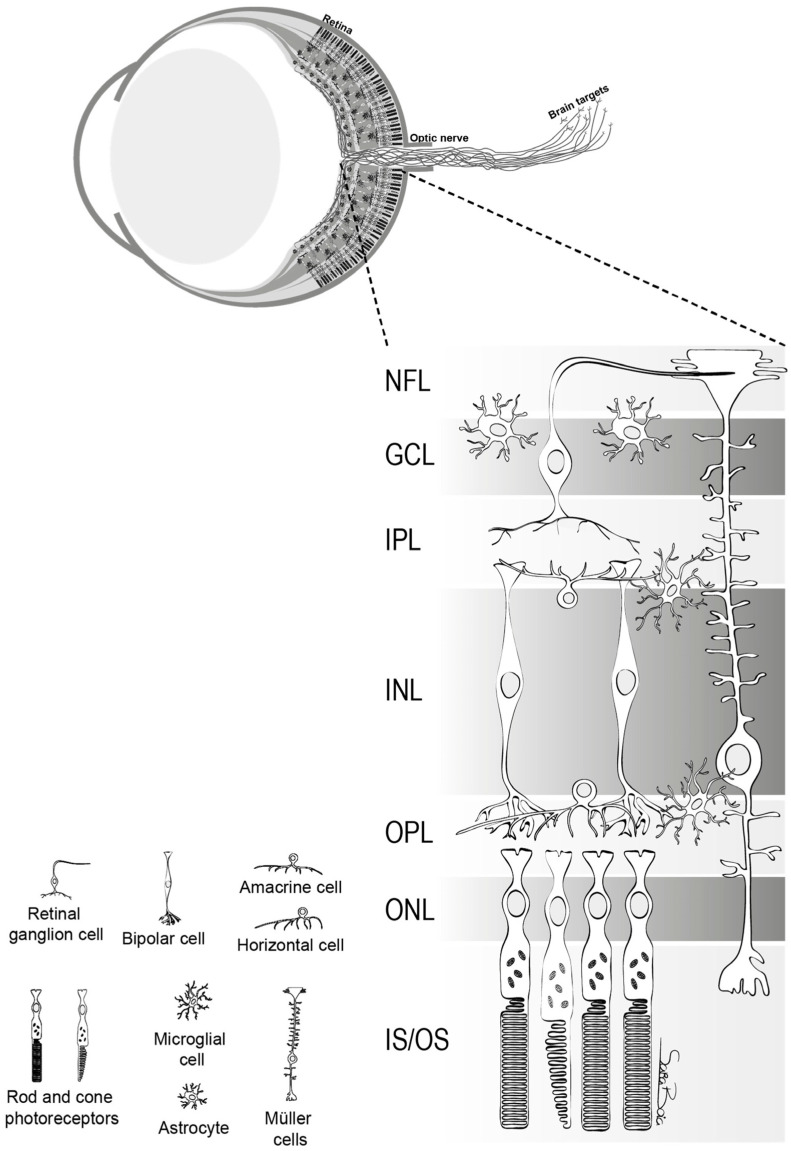
RGCs play a crucial role in transmitting visual stimuli from the retina to the brain. From a morphological standpoint, RGCs consist of a cellular body, a complex dendritic structure, and a single axon. The primary function of dendrites is to receive synaptic inputs from bipolar and amacrine cells, which are intermediary neurons responsible for acquiring signals generated by photoreceptors [5,6,7] (Figure 1). The presence of hierarchical circuitry allows RGCs to engage in intricate signal integration and processing, leading to the conversion of photoreceptor responses into action potentials. The action potentials propagate through the RGC axons, which together constitute the optic nerve. They travel through the optic disc and optic canal to establish synaptic connections in the lateral geniculate nucleus (LGN), located in the thalamus [6]. The visual information is transmitted from the LGN to the primary visual cortex (V1) for the purpose of undergoing higher-level processing and integration. Therefore, RGCs play a crucial role in connecting the initial light detection in the retina to the following brain visual perception.
RGC는
망막에서 뇌로 시각 자극을 전달하는 데 중요한 역할을 합니다.
형태학적인 관점에서 볼 때
RGC는
세포체,
복잡한 수상돌기 구조 및 단일 축삭으로 구성됩니다.
수상 돌기의 주요 기능은
광수용체에서 생성된 신호를 획득하는
중간 뉴런인 양극성 및
아마크린 세포로부터 시냅스 입력을 받는 것입니다[5,6,7](그림 1).
계층적 회로의 존재는 RGC가 복잡한 신호 통합 및 처리에 관여하여 광수용체 반응을 활동 전위로 전환할 수 있도록 합니다. 활동 전위는 시신경을 구성하는 RGC 축삭돌기를 통해 전파됩니다. 이들은 시신경 디스크와 시도를 통해 이동하여 시상에 위치한 측생식핵(LGN)에서 시냅스 연결을 형성합니다[6]. 시각 정보는 더 높은 수준의 처리와 통합을 위해 LGN에서 일차 시각 피질(V1)로 전송됩니다. 따라서 RGC는 망막의 초기 빛 감지를 다음 단계의 뇌 시각 지각으로 연결하는 데 중요한 역할을 합니다.
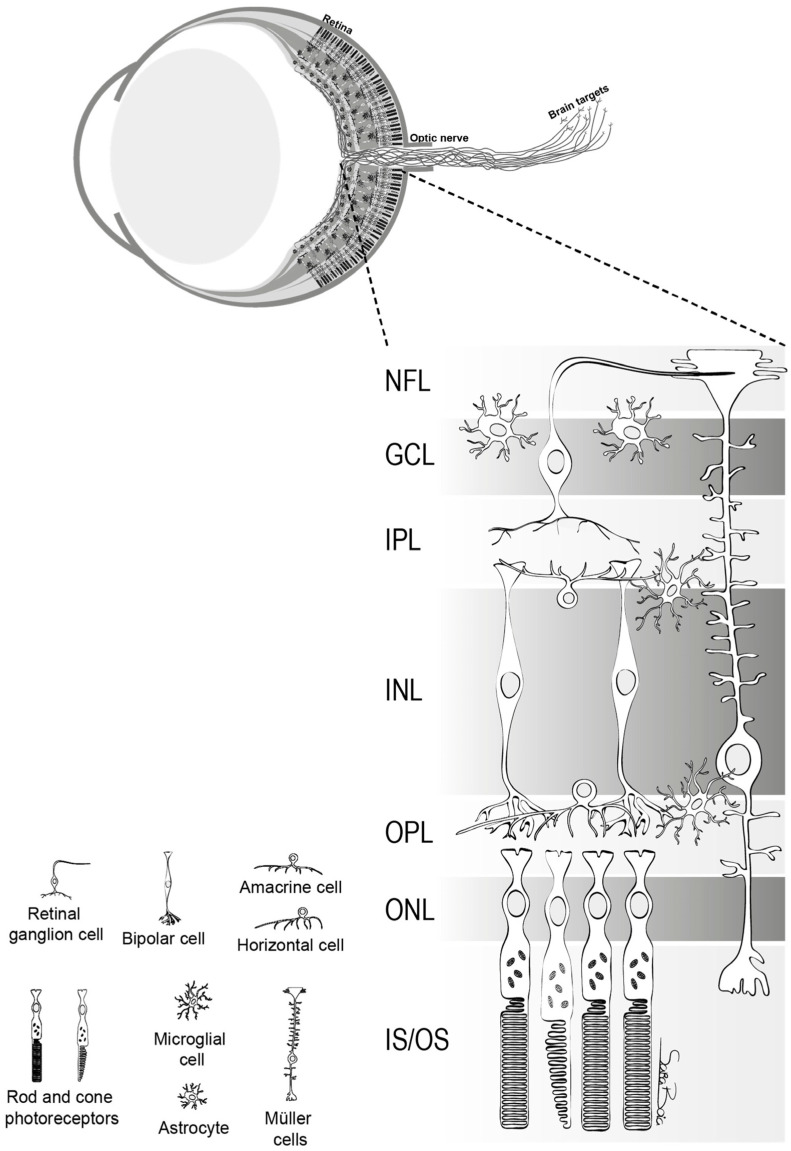
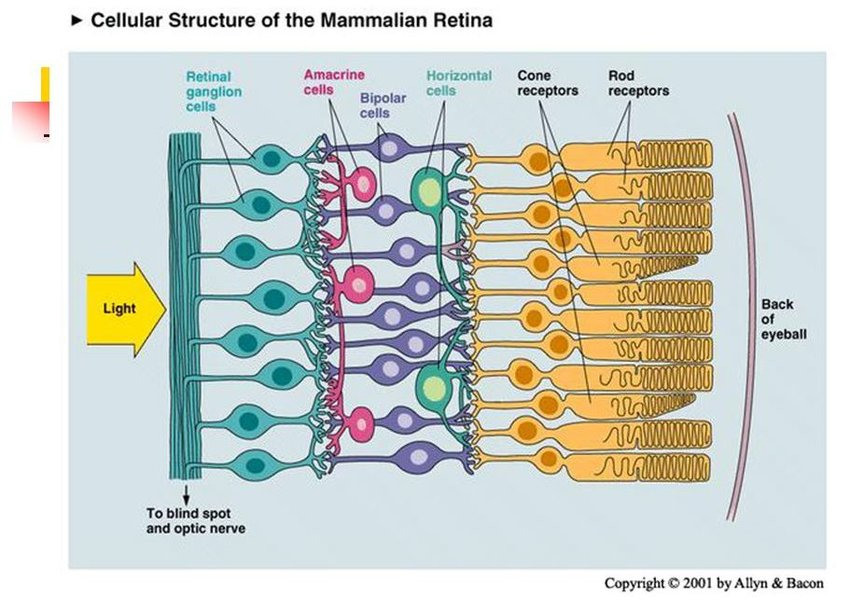
The structure of retina. This figure delineates the stratified architecture of the neural sensory retina, illustrating the cellular arrangement using distinct nuclear and plexiform layers. Photoreceptor nuclei, specifically those of rods and cones, reside within the outer nuclear layer (ONL), while the nuclei of various interneurons, including amacrine, bipolar, and horizontal cells, are primarily situated in the inner nuclear layer (INL). Retinal ganglion cells (RGCs) inhabit the ganglion cell layer (GCL), with their axonal projections extending through the nerve fiber layer (NFL). The retina comprises two macroglial variants: Müller cells, traversing the entire retinal thickness, and astrocytes, localized within the GCL. Microglia are chiefly found in the inner retinal regions and the outer plexiform layer (OPL) [5].
망막의 구조.
이 그림은 신경 감각 망막의 계층화된 구조를 묘사한 것으로, 뚜렷한 핵층과 플렉시폼 층을 사용하여 세포 배열을 보여줍니다. 광수용체 핵, 특히 막대와 원뿔의 핵은 외핵층(ONL)에 위치하며, 아마크린, 양극성, 수평 세포를 포함한 다양한 내부 뉴런의 핵은 주로 내핵층(INL)에 위치합니다. 망막 신경절 세포(RGC)는 신경절 세포층(GCL)에 서식하며 축삭 돌기가 신경 섬유층(NFL)을 통해 확장되어 있습니다. 망막은 두 개의 대교세포 변종으로 구성됩니다: 망막 전체 두께를 가로지르는 뮐러 세포와 GCL 내에 국한된 성상교세포가 그것입니다. 미세아교세포는 주로 망막 안쪽 영역과 바깥쪽 플렉시폼 층(OPL)에서 발견됩니다[5].
In the human retina, a variety of RGCs are present, each specializing in distinct visual functions. The presence of heterogeneity among RGCs is a fundamental quality, as these cells can be differentiated into multiple subtypes based on their distinct morphological and physiological characteristics. Midget RGCs (P-Cell), comprising approximately 80% of RGCs, are primarily associated with the processing of visual information at a high level of detail and the discriminating of colors [8]. Parasol RGCs (M-Cell), distinct from midget cells, are known to play a role in the detection of motion and changes in luminance [9]. Midget RGCs receive inputs primarily from single cone photoreceptors and exhibit a sustained response to stimuli; the axons of midget RGCs project to dorsal parvocellular layers of the LGN. Parasol RGCs exhibit a larger dendritic field compared to midget RGCs and have a transient response to visual stimuli; the axons of parasol RGCs project to the ventral magnocellular layers of the RGC [10]. Small bistratified RGCs, which constitute about 5–8% of the total, are an additional prominent subtype that receives synaptic inputs from both ON and OFF bipolar cells, allowing them to integrate signals from both pathways and contribute to the encoding of color contrast, particularly along the blue–yellow axis. These cells are components of the koniocellular pathway, implicated in color processing and integrating diverse visual features [8,11]. Smooth monostratified RGCs, divided into ON and OFF cells, can relay spatial information in a complex manner [12]. Melanopsin-containing intrinsically photosensitive RGCs (ipRGCs), essential for non-visual functions, represent about 1% of the total RGC population [13]. Additionally, there are miscellaneous RGCs that defy existing classifications, underscoring the retina’s intricate complexity [14]. Each specific subtype of RGCs makes distinct contributions to many areas of visual perception, encompassing spatial resolution, contrast sensitivity, and chromatic discrimination [15,16]. Furthermore, it is important to note that certain subtypes of RGCs exhibit varying levels of vulnerability to degenerative alterations in glaucomatous circumstances. For example, in a pressure-induced environment, midget and parasol cells undergo degeneration that begins with dendritic arbor and concludes with cell soma shrinkage [10]. As a result, it is crucial to develop a comprehensive understanding of the distinct functions and contributions of these subtypes.
인간의 망막에는
각각 고유한 시각 기능에 특화된 다양한 RGC가 존재합니다.
이러한 세포는
뚜렷한 형태적 및 생리적 특성에 따라
여러 하위 유형으로 분화될 수 있기 때문에
RGC 간의 이질성은 근본적인 특성입니다.
RGC의 약 80%를 차지하는 미젯 RGC(P-Cell)는
주로 높은 수준의 시각 정보 처리 및 색상 식별과 관련이 있습니다[8].
난쟁이 세포와 구별되는 파라솔 RGC(M-Cell)는
움직임과 휘도의 변화를 감지하는 역할을 하는 것으로 알려져 있습니다[9]. 난쟁이 RGC는 주로 단일 원추형 광수용체로부터 입력을 받고 자극에 대한 지속적인 반응을 나타내며, 난쟁이 RGC의 축삭은 LGN의 등쪽 부세포층으로 돌출합니다.
파라솔 RGC는
난쟁이 RGC에 비해 더 큰 수상돌기장을 나타내며
시각 자극에 일시적으로 반응하고,
파라솔 RGC의 축삭은 RGC의 복부 자세포층으로 돌출됩니다 [10]. 전체의 약 5-8%를 차지하는 작은 이층화 RGC는 ON 및 OFF 양극성 세포로부터 시냅스 입력을 받는 또 다른 두드러진 하위 유형으로, 두 경로의 신호를 통합하고 특히 청황색 축을 따라 색 대비의 인코딩에 기여할 수 있습니다. 이 세포는 색 처리와 다양한 시각적 특징 통합에 관여하는 코니오세포 경로의 구성 요소입니다[8,11].
켜짐과 꺼짐 세포로 나뉘는 매끄러운 단일 층화 RGC는 복잡한 방식으로 공간 정보를 전달할 수 있습니다 [12]. 비시각적 기능에 필수적인 멜라놉신 함유 본질적으로 광과민성 RGC(ipRGC)는 전체 RGC 개체군의 약 1%를 차지합니다[13].
또한
망막의 복잡한 복잡성을 강조하는 기존의 분류를 무시하는
기타 RGC도 존재합니다 [14].
RGC의 각 특정 하위 유형은
공간 해상도, 대비 감도 및 색채 식별을 포함하여
시각 인식의 여러 영역에 뚜렷한 기여를 합니다 [15,16].
또한, 녹내장 상황에서
특정 하위 유형의 RGC가 퇴행성 변화에 대해
다양한 수준의 취약성을 보인다는 점에 유의하는 것이 중요합니다.
예를 들어, 압력 유발 환경에서 난쟁이 세포와 파라솔 세포는 수지상 정으로 시작하여 세포 체질 수축으로 끝나는 퇴행을 겪습니다 [10]. 따라서 이러한 하위 유형의 고유한 기능과 기여에 대한 포괄적인 이해가 중요합니다.
3. Pathophysiology of Glaucoma and Current Theories on Retinal Ganglion Cell Vulnerability and Functionality in Glaucoma
Glaucoma is a heterogeneous group of ocular disorders that culminates in RGC degeneration and visual field loss. The mechanism underlying glaucomatous optic neuropathy is multifaceted, with theories on RGC vulnerability that include mechanical, vascular, excitatory, neurotrophic factor deprivation, and immune and inflammation mechanisms (Table 1) [17].
녹내장의 병태생리 및 녹내장의 망막 신경절 세포 취약성 및 기능에 대한 최신 이론
녹내장은
망막 신경절 세포의 변성과
시야 손실로 정점에 이르는 이질적인 안과 질환 그룹입니다.
녹내장성 시신경병증의 기저에 있는 메커니즘은
다면적인데,
기계적,
혈관,
흥분성,
신경 영양 인자 결핍, 면
역 및 염증 메커니즘을 포함하는 RGC 취약성에 대한 이론이 있습니다(표 1)[17].
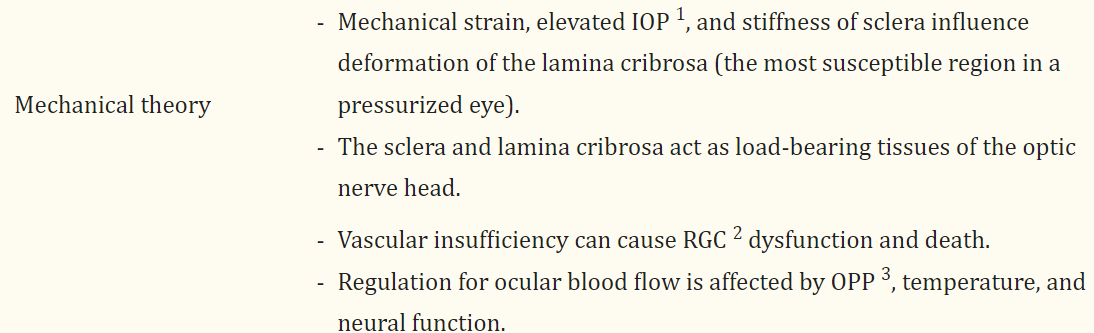


Table 1
Overview of glaucoma pathophysiology and contemporary theoretical frameworks.
TheoriesKey Points
| Mechanical theory |
|
| Vascular theory |
|
| Excitotoxicity theory |
|
| Neurotrophic factor deprivation theory |
|
| Oxidative Stress and Inflammation |
|
1 IOP, intraocular pressure; 2 RGC, retinal ganglion cell; 3 OPP, ocular perfusion pressure; 4 NMDA, N-methyl-d-aspartate; 5 BDNF, brain-derived neurotrophic factor; 6 TrkB, tropomyosin receptor kinase B; 7 CNTF, ciliary neurotrophic factor; 8 NGF, nerve growth factor; 9 GDNF, glial cell line–derived neurotrophic factor; 10 ROS, reactive oxygen species; 11 HSP, heat shock protein.
3.1. Mechanical Theories
The pathophysiology of glaucoma is complex, involving both mechanical and vascular components. Mechanical strain and compressive forces are exerted at the lamina cribrosa, recognized as the most susceptible region of a pressurized eye, subsequently affecting the structural stability and functionality of RGC axons [18]. Research showed that elevated IOP from 5 to 50 mmHg over 24 h resulted in an average posterior deformation of the central lamina measuring 79 μm in human donor eyes and posterior displacement of the central lamina of 12 μm in response to an acute increase of IOP from 10 to 25 mmHg in human eyes [19,20]. In addition, connective tissue of the peripapillary sclera and scleral canal wall are also responsible for bearing the forces generated by the IOP. The structural stiffness of the sclera plays a significant role in determining the deformation of the lamina cribrosa. Compliant sclera enables expansion of the scleral canal in response to elevated IOP, which results in tightening of the laminar beams within the canal and increases lamina resistance against posterior deformation. On the other hand, a rigid sclera exhibits limited capacity for canal expansion, hence necessitating the sole reliance on the structural rigidity of the lamina cribrosa to withstand the stress associated with elevated IOP [20]. Therefore, the sclera and lamina cribrosa serve as the load-bearing tissues of the optic nerve head [21].
녹내장의 병태생리는
기계적 요소와 혈관적 요소가 모두 관여하는 복잡한 구조입니다.
안압이 높은 눈에서 가장 취약한 부위로 알려진 섬유층에
기계적 변형과 압축력이 가해져
RGC 축삭의 구조적 안정성과 기능에 영향을 미칩니다[18].
연구에 따르면 24시간 동안 안압이
5에서 50mmHg로 상승하면
인간 기증자 눈에서 평균 79μm의 중심층 후방 변형이 발생하고,
인간 눈에서 안압이 10에서 25mmHg로 급격히 상승하면
중심층이 12μm 후방으로 변위되는 것으로 나타났습니다 [19,20].
또한
유두 주변 공막과 공막관 벽의 결합 조직도
안압에 의해 생성되는 힘을 견디는 역할을 합니다.
공막의 구조적 강성은 층막의 변형을 결정하는 데 중요한 역할을 합니다. 공막이 유연하면 안압 상승에 반응하여 공막이 확장되고, 그 결과 공막 내 층류 빔이 조여져 후방 변형에 대한 층류 저항이 증가합니다. 반면, 경직된 공막은 안압 상승과 관련된 스트레스를 견디기 위해 공막의 구조적 강성에 전적으로 의존해야 하므로 공막의 확장 능력이 제한적입니다[20]. 따라서 공막과 섬유층은 시신경 머리의 하중을 견디는 조직으로 작용합니다[21].
3.2. Vascular Theories
The aforementioned discussion on mechanical theory suggests axonal compression at the lamina cribrosa, hindrance of axoplasmic flow, and disruption of retrograde neurotrophin transport to RGCs, ultimately resulting in cellular demise. Another aspect of RGC vulnerability is vascular insufficiency, where it is hypothesized that impaired perfusion pressure, disruption of vascular autoregulation, and loss of neurovascular coupling can lead to RGC dysfunction and death. In a study conducted by Shiga et al., the results of laser flow speckle flowgraphy indicated a statistically significant decrease in ocular blood flow in patients with preperimetric glaucoma compared to normal subjects [22].
앞서 언급한 기계적 이론에 대한 논의는
층상 돌기에서의
축삭 압박,
축삭질 흐름의 방해,
RGC로의 역행성 뉴로트로핀 수송의 중단이
궁극적으로 세포 사멸을 초래한다는 것을 시사합니다.
RGC 취약성의 또 다른 측면은
혈관 기능 부전으로,
관류 압력 장애,
혈관 자가 조절 장애,
신경 혈관 결합 상실이 RGC 기능 장애와 사망으로 이어질 수 있다는 가설이 있습니다.
Shiga 등이 수행한 연구에서 레이저 흐름 얼룩 흐름 그래피 결과는 전녹내장 환자의 안구 혈류가 정상인에 비해 통계적으로 유의하게 감소하는 것으로 나타났습니다[22].
Ocular perfusion pressure (OPP) is defined as the sum of the systolic arterial blood pressure and one third of the difference between the systolic and diastolic pressures, minus the IOP. OPP signifies the oxygen supply and blood flow to the optic nerve; thus, it has been hypothesized that a reduction in OPP could increase the optic disc’s susceptibility to damage, thereby increasing the likelihood of glaucoma progression or development [23]. Many studies have revealed a positive correlation between a reduction in OPP and an elevated prevalence of open-angle glaucoma [24,25,26]. Additionally, previous research utilizing microspheres indicated a specific reduction of volume flow within the prelaminar and anterior laminar capillary beds when the OPP fell below 30 mm Hg [27,28]. Regulation of ocular blood flow is affected by OPP, temperature, and neural function, and blood flow to the optic nerve head is mainly regulated by endothelial cells and circulating hormones [29].
안구 관류 압력(OPP)은
수축기 동맥 혈압과 수축기 및 이완기 혈압 차이의 1/3에서
안압을 뺀 값의 합으로 정의됩니다.
OPP는
시신경에 대한 산소 공급과 혈류를 의미하므로,
OPP가 감소하면 시신경이 손상되기 쉬워
녹내장 진행 또는 발병 가능성이 높아진다는 가설이 제기되었습니다 [23].
많은 연구에서
OPP 감소와
개방각 녹내장 유병률 증가 사이에
긍정적인 상관관계가 있음이 밝혀졌습니다 [24,25,26].
또한, 마이크로스피어를 활용한 이전 연구에서는 OPP가 30mmHg 이하로 떨어질 때 전층 및 전층 모세혈관층 내의 체적 흐름이 구체적으로 감소하는 것으로 나타났습니다[27,28]. 안구 혈류의 조절은 OPP, 온도 및 신경 기능의 영향을 받으며, 시신경두로의 혈류는 주로 내피 세포와 순환 호르몬에 의해 조절됩니다 [29].
Vascular autoregulation refers to the regulatory mechanism that counteracts fluctuations in perfusion pressure, functioning optimally within a specific range of perfusion pressure. This mechanism can be compromised in conditions like glaucoma as well as other diseases, including diabetes mellitus. Various factors are involved in autoregulation, such as metabolic, myogenic, and neurogenic components. Direct autoregulation of ocular blood flow is difficult to assess, and many techniques have been used to measure ocular circulation, such as color Doppler imaging, scanning laser Doppler flowmeters, and optical coherence tomography (OCT). A study revealed individuals with glaucoma exhibited inadequate compensatory reaction of flow velocities in the short posterior ciliary artery (SPCA) to alterations in body position compared with healthy individuals, suggesting inadequate autoregulatory control may serve as a contributing factor to glaucoma pathogenesis [30].
혈관 자가 조절은
관류 압력의 변동에 대응하는 조절 메커니즘으로,
특정 범위의 관류 압력 내에서 최적으로 작동합니다.
이 메커니즘은 녹내장과 같은 질환뿐만 아니라
당뇨병을 포함한 다른 질환에서도 손상될 수 있습니다.
자가 조절에는 대사성, 근육성 및 신경성 요소와 같은 다양한 요인이 관여합니다. 안구 혈류의 직접적인 자가 조절은 평가하기 어렵기 때문에 컬러 도플러 영상, 스캐닝 레이저 도플러 유량계, 빛간섭단층촬영(OCT) 등 안구 순환을 측정하는 데 많은 기술이 사용되어 왔습니다. 한 연구에 따르면 녹내장 환자는 건강한 사람에 비해 신체 위치 변화에 대한 짧은 후방 섬모 동맥(SPCA)의 유속 보상 반응이 불충분한 것으로 나타났는데, 이는 부적절한 자가 조절 조절이 녹내장 발병의 원인으로 작용할 수 있음을 시사합니다[30].
The central nervous system has a strong correlation between neuronal activity and blood flow, indicating a high level of coordination. Neurovascular coupling is the phenomenon wherein heightened neural activity leads to a concomitant augmentation in blood flow to the appropriate region [31]. A study demonstrated a lower cerebral blood flow and functional connectivity strength coupling in glaucoma patients compared to controls, and the reduced ratio was significantly correlated with visual field defects and glaucoma stage, suggesting impaired neurovascular coupling in glaucoma patients [32].
중추신경계는 신경세포 활동과 혈류 사이에 강한 상관관계가 있으며, 이는 높은 수준의 조정을 나타냅니다. 신경 혈관 결합은 신경 활동이 증가하면 해당 부위로의 혈류가 수반적으로 증가하는 현상입니다 [31]. 한 연구에 따르면 녹내장 환자는 대조군에 비해 뇌 혈류와 기능적 연결 강도 결합이 낮았으며, 감소된 비율은 시야 결손 및 녹내장 단계와 유의미한 상관관계가 있어 녹내장 환자의 신경혈관 결합이 손상된 것으로 나타났습니다 [32].
Numerous mediators orchestrate the regulation of ocular blood flow in glaucoma, which is a critical determinant of the health of the RGC and optic nerve head. Central among these mediators is endothelin-1 (ET-1), a potent vasoconstrictive peptide, which is elevated in aqueous and plasma concentrations of glaucoma patients [33,34,35,36,37]. Clinical studies revealed worsening visual fields have higher plasma ET-1 compared to normal visual fields [35], and animal studies demonstrated that low doses of ET-1 can induce glaucomatous changes in primates [38]. Increased ET-1 levels result in an increase in IOP, which decreases ocular blood flow and astrocyte proliferation; consequently, this may ultimately lead to the degeneration of RGCs [39]. A recent study utilized advanced imaging techniques, specifically OCT and OCT angiography, to assess vessel density in the peripapillary and macular regions of patients with glaucoma, revealing an inverse relationship with systemic ET-1 concentrations [37]. Multivariate analyses indicated that IOP plays a less predictive role in the diminishment of retinal blood flow when compared to ET-1 levels in a glaucoma context. Elevated peripheral ET-1 has been identified as a potential risk indicator for monitoring vascular alterations in the optic nerve head of eyes affected by glaucoma. Conversely, nitric oxide acts as a pivotal vasodilator by stimulating cyclic guanine monophosphate (cGMP) to reduce intracellular calcium and thereby relaxation of smooth muscle cells and pericytes [40]. In glaucoma patients, nitric oxide availability is reduced, which can cause a disruption in the equilibrium between vasoconstriction and vasodilation and ultimately results in a reduction of blood flow to the optic nerve head [41].
녹내장에서는 수많은 매개체가 안구 혈류의 조절을 조율하며, 이는 RGC와 시신경 머리의 건강을 결정하는 중요한 요인입니다. 이러한 매개체 중 중심에는 강력한 혈관 수축 펩타이드인 엔도텔린-1(ET-1)이 있으며, 녹내장 환자의 수성 및 혈장 농도가 증가합니다[33,34,35,36,37]. 임상 연구에 따르면 악화된 시야는 정상 시야에 비해 혈장 ET-1이 더 높으며[35], 동물 연구에서는 저용량의 ET-1이 영장류에서 녹내장성 변화를 유발할 수 있다는 것이 입증되었습니다[38]. ET-1 수치가 증가하면 안압이 증가하여 안구 혈류와 성상세포 증식이 감소하고, 결과적으로 이는 궁극적으로 RGC의 변성을 초래할 수 있습니다 [39]. 최근의 한 연구에서는 녹내장 환자의 유두 주변 및 황반 부위의 혈관 밀도를 평가하기 위해 첨단 영상 기술, 특히 OCT 및 OCT 혈관 조영술을 활용하여 전신 ET-1 농도와 역의 관계를 밝혀냈습니다 [37]. 다변량 분석에 따르면 녹내장 맥락에서 안압은 ET-1 수치에 비해 망막 혈류의 감소를 예측하는 데 덜 중요한 역할을 하는 것으로 나타났습니다. 말초 ET-1의 상승은 녹내장의 영향을 받는 눈의 시신경 머리에서 혈관 변화를 모니터링하는 잠재적 위험 지표로 확인되었습니다. 반대로 산화질소는 사이클릭 구아닌 모노포스페이트(cGMP)를 자극하여 세포 내 칼슘을 감소시켜 평활근 세포와 주변 세포의 이완을 유도함으로써 중추적인 혈관 확장제 역할을 합니다[40]. 녹내장 환자의 경우 산화질소 가용성이 감소하여 혈관 수축과 혈관 확장 사이의 균형이 깨지고 궁극적으로 시신경 머리로의 혈류가 감소할 수 있습니다 [41].
3.3. Excitotoxicity Theory
Research into the role of excitotoxicity, which is mediated by excessive glutamate release and subsequent overstimulation of N-methyl-d-aspartate (NMDA) receptors on RGCs, has revealed the molecular and cellular pathways that contribute to RGC injury in glaucoma. Glutamate serves as a primary neurotransmitter inside the retinal network, facilitating communication between the photoreceptors, bipolar cells, and RGCs [42]. The process of glutamate neurotransmission is intricately regulated by inhibitory and modulatory neurotransmitters, including γ-aminobutyric acid (GABA), glycine, acetylcholine, dopamine, serine, substance P, and several neuropeptides [43]. Excessive glutamate release causes overactivation of the NMDA receptor, which induces calcium influx, oxidative stress, and mitochondrial dysfunction, all of which contribute to the degeneration of RGCs. An increase in glutamate has not been consistently seen in retinal pathologies such as glaucoma [44]; however, due to the progressive nature of glaucoma, glutamate levels may not significantly increase acutely or accumulate during the course of the disease and may only rise in specific parts of the retina or optic nerve.
과도한 글루타메이트 방출과
그에 따른 RGC의 N-메틸-D-아스파르트산염(NMDA) 수용체의 과자극에 의해 매개되는
흥분 독성의 역할에 대한 연구를 통해 녹
내장에서 RGC 손상에 기여하는 분자 및 세포 경로가 밝혀졌습니다.
글루타메이트는
망막 네트워크 내에서 주요 신경전달물질로 작용하여
광수용체,
양극성 세포,
RGC 간의 통신을 촉진합니다[42].
글루타메이트 신경 전달 과정은
γ- 아미노부티르산(GABA),
글리신,
아세틸콜린,
도파민,
세린,
물질 P 및 여러 신경 펩타이드를 포함한
억제 및 조절 신경 전달 물질에 의해 복잡하게 조절됩니다 [43].
과도한 글루타메이트 방출은
NMDA 수용체의 과활성화를 유발하여
칼슘 유입,
산화 스트레스 및 미토콘드리아 기능 장애를 유도하며,
이 모든 것이 RGC의 퇴화에 기여합니다.
녹내장과 같은 망막 병리에서 글루타메이트의 증가는 일관되게 관찰되지 않았지만[44], 녹내장의 진행성 특성으로 인해 글루타메이트 수치가 급격히 증가하거나 질병이 진행되는 동안 축적되지 않고 망막이나 시신경의 특정 부분에서만 상승할 수 있습니다.
Generally, when glutamate is released, the initial depolarization of the cell membrane through α-amino-3-hydroxy-5-methylisoxazole-4-propionate (AMPA) receptor activation leads to an elevation in the likelihood of NMDA receptor opening. This triggers a transient influx of calcium ions into the intracellular space. Calcium can also be released via the inositol 1,4,5-trisphosphate receptors (IP3R) and the ryanodine receptors (RyR) that are present on the membrane of the endoplasmic reticulum. Functional mitochondrial metabolism plays a crucial role in maintaining intracellular calcium homeostasis through the reuptake of calcium by the Sarco-Endoplasmic Reticulum Calcium ATPase (SERCA) and the extrusion of calcium to the extracellular space via the sodium-calcium ATPase. Furthermore, the extrusion of calcium is facilitated by the sodium-calcium exchanger (NCX) through the utilization of the physiological sodium gradient. The failure of RGCs to adequately regulate or remove excessive calcium ions disrupts the homeostatic mechanisms, which ultimately culminates in cellular demise. The involvement of NCX in the process of retinal cell death triggered by NMDA and ischemia-reperfusion has been suggested [45,46]. High levels of intracellular calcium along with heightened activity of catabolic enzymes can trigger a cascade of events, leading to RGC apoptosis and necrosis [47,48]. The subsequent cascade encompasses mitochondrial membrane depolarization, activation of caspases, generation of harmful oxygen and nitrogen free radicals, and manifestation of cellular toxicity.
일반적으로
글루타메이트가 방출되면
α-아미노-3-하이드록시-5-메틸이소옥사졸-4-프로피오네이트(AMPA) 수용체 활성화를 통한
세포막의 초기 탈분극으로 인해 NMDA 수용체 개방 가능성이 높아지게 됩니다.
이는 칼슘 이온이 세포 내 공간으로 일시적으로 유입되는 것을 촉발합니다.
칼슘은 소포체 막에 존재하는 이노시톨 1,4,5-트리스포스페이트 수용체(IP3R)와 라이노딘 수용체(RyR)를 통해서도 방출될 수 있습니다. 기능적 미토콘드리아 대사는 소포체 칼슘 ATPase(SERCA)에 의한 칼슘 재흡수와 나트륨-칼슘 ATPase를 통한 칼슘의 세포 외 공간으로의 배출을 통해 세포 내 칼슘 항상성을 유지하는 데 중요한 역할을 합니다. 또한 생리적 나트륨 구배를 활용하여 나트륨-칼슘 교환기(NCX)에 의해 칼슘의 압출이 촉진됩니다. RGC가 과도한 칼슘 이온을 적절히 조절하거나 제거하지 못하면 항상성 메커니즘이 파괴되어 궁극적으로 세포 사멸에 이르게 됩니다. NMDA와 허혈 재관류에 의해 촉발되는 망막 세포 사멸 과정에서 NCX의 관여가 제안되었습니다 [45,46]. 세포 내 칼슘 수치가 높고 이화 효소의 활성이 높아지면 일련의 사건이 촉발되어 RGC 세포 사멸과 괴사로 이어질 수 있습니다 [47,48]. 후속 캐스케이드에는 미토콘드리아 막 탈분극, 카스파제 활성화, 유해 산소 및 질소 유리기 생성, 세포 독성 발현이 포함됩니다.
Normal glutamate excitotoxicity is minimized by its quick absorption by glutamate transporters in glial cells surrounding RGCs. The majority of glutamate transporters in the retina are located at the synaptic cleft in order to quickly remove released glutamate and prevent overflow [49]. Glutamate uptake by Müller cells via glutamate/aspartate transporters (GLAST or EAAT1) is crucial for maintaining physiological glutamate levels [50]. Retinal glia and neurons produce several glutamate transporters, such as GLT-1 (EAAT2), excitatory amino acid carrier 1 (EAAC1 in rats or EAAT3 in humans), EAAT4, and EAAT5 in rats. Decreased retinal GLAST has been correlated with glaucoma in rats and mice [51,52], and GLAST-deficient mice displayed loss of RGCs [53].
The profound vulnerability of RGCs is underscored by the imbalance between neurotransmitter release and uptake, as well as the cascade of intracellular events triggered by excessive receptor stimulation.
정상적인 글루타메이트 흥분 독성은
RGC를 둘러싼 신경교세포의 글루타메이트 수송체에 의해 빠르게 흡수되어 최소화됩니다.
망막의 글루타메이트 수송체의 대부분은 시냅스 틈새에 위치하여 방출된 글루타메이트를 빠르게 제거하고 과잉을 방지합니다 [49]. 글루타메이트/아스파르트산염 수송체(GLAST 또는 EAAT1)를 통한 뮐러 세포의 글루타메이트 흡수는 생리적 글루타메이트 수준을 유지하는 데 매우 중요합니다 [50]. 망막 신경교세포와 뉴런은 GLT-1(EAAT2), 흥분성 아미노산 운반체 1(쥐의 경우 EAAC1, 인간의 경우 EAAT3), EAAT4 및 EAAT5와 같은 여러 글루타메이트 수송체를 생성합니다. 망막 GLAST 감소는 쥐와 마우스에서 녹내장과 상관관계가 있으며[51,52], GLAST 결핍 마우스는 RGC가 소실된 것으로 나타났습니다[53].
RGC의 심각한 취약성은 신경전달물질 방출과 흡수 사이의 불균형과 과도한 수용체 자극에 의해 촉발되는 일련의 세포 내 사건으로 인해 강조됩니다.
3.4. Neurotrophic Factor Deprivation Theory
According to the neurotrophic factor deprivation theory, the interruption of axonal transport results in a diminished supply of neurotrophic factors, which are essential for the survival of RGCs. Neurotrophins, which are diffusible trophic molecules, enhance the survival of mature neurons in the central nervous system (CNS), which degenerate in response to an extensive range of stimuli. Brain-derived neurotrophic factor (BDNF) has garnered significant interest within the field of neurotrophins due to its influential impact on the survival of RGCs. The initial evidence of BDNF’s neuroprotective properties in RGCs occurred in 1986 [54]. BDNF is produced locally in the retina by RGCs and glia, but it is also strongly expressed in the superior colliculus, where it is retrogradely transported to the optic nerve head and to the cell bodies of RGCs [49,55]. The effects of BDNF are mediated through tropomyosin receptor kinase B (TrkB) on the presynaptic cell surface, and TrkB activation leads to three major signaling cascades (mitogen-activated protein kinase (MAPK), phosphatidylinositol 3-kinase (PI3K), and phospholipase Cγ (PLC-γ)), which are responsible for promoting the survival of RGCs [56]. Elevated IOP and the resulting mechanical strain on the optic nerve head may hinder the retrograde transport of BDNF in glaucoma. When the stressed RGC is deprived of BDNF, Jun-N terminal kinase (JNK)-mediated signaling may be triggered, which may activate the proapoptotic BCL-2 family of proteins and cause mitochondrial dysfunction [55,57,58]. Numerous studies have demonstrated a correlation between RGC loss and impaired BDNF-TrkB signaling [59,60,61]. Consistent clinically, BDNF deficit was demonstrated in serum, aqueous humor, and lacrimal tears of patients with early glaucomatous change [62,63]. Apart from BDNF, additional neurotrophic factors have been linked to RGC health, including ciliary neurotrophic factor (CNTF), nerve growth factor (NGF), and glial cell line–derived neurotrophic factor (GDNF) [57].
The multifaceted nature of neurotrophic factors in the retina emphasizes the importance of its support in RGC health.
신경 영양 인자 결핍 이론에 따르면,
축삭 수송이 중단되면
RGC의 생존에 필수적인 신경 영양 인자의 공급이 감소합니다.
확산 가능한 영양 분자인 뉴로트로핀은
광범위한 자극에 반응하여 퇴화하는 중추신경계(CNS)에서 성숙한 뉴런의 생존을 향상시킵니다.
뇌유래신경영양인자(BDNF)는
RGC의 생존에 미치는 영향력 때문에
뉴로트로핀 분야에서 큰 관심을 받고 있습니다.
RGC에서 BDNF의 신경 보호 특성에 대한 최초의 증거는 1986년에 나왔습니다[54]. BDNF는 RGC와 신경교세포에 의해 망막에서 국소적으로 생성되지만, 시신경 머리와 RGC의 세포체로 역행적으로 수송되는 상부 컬리큘러스에서도 강하게 발현됩니다 [49,55]. BDNF의 효과는 시냅스 전 세포 표면의 트로포마이오신 수용체 키나아제 B(TrkB)를 통해 매개되며, TrkB 활성화는 RGC의 생존을 촉진하는 세 가지 주요 신호 캐스케이드(미토겐 활성화 단백질 키나제(MAPK), 포스파티딜이노시톨 3-kinase(PI3K), 포스포리파제 Cγ(PLC-γ))로 이어집니다 [56]. 안압 상승과 이로 인한 시신경 머리의 기계적 긴장은 녹내장에서 BDNF의 역행 수송을 방해할 수 있습니다. 스트레스를 받은 RGC에 BDNF가 부족하면 Jun-N 말단 키나아제(JNK)를 매개로 한 신호가 유발되어 세포 자멸사 유도 BCL-2 단백질 계열을 활성화하고 미토콘드리아 기능 장애를 일으킬 수 있습니다 [55,57,58]. 수많은 연구에서 RGC 손실과 BDNF-TrkB 신호 전달 장애 사이의 상관관계가 입증되었습니다 [59,60,61]. 임상적으로 일관되게 초기 녹내장 변화 환자의 혈청, 수성액, 눈물샘에서 BDNF 결핍이 입증되었습니다[62,63]. BDNF 외에도 섬모 신경 영양 인자(CNTF), 신경 성장 인자(NGF), 신경 아교 세포주 유래 신경 영양 인자(GDNF) 등 추가적인 신경 영양 인자가 RGC 건강과 연관되어 있습니다[57].
망막에서 신경 영양 인자의 다면적인 특성은 망막 건강에서 신경 영양 인자의 지원의 중요성을 강조합니다.
3.5. Oxidative Stress and Inflammation
Oxidative stress and neuroinflammation are implicated in the pathogenesis of glaucoma, where reactive oxygen species (ROS) and inflammatory mediators precipitate cellular damage and contribute to an excitotoxic environment, thereby exacerbating RGC death. Multiple factors can contribute to increased ROS production in glaucomatous eyes. Ischemia-reperfusion injury, elevated IOP, and mitochondrial dysfunction are among the principal factors that contribute to the overproduction of ROS in the optic nerve head and retina [49,64]. The principal roles of mitochondria encompass the synthesis of adenosine triphosphate (ATP) via the oxidative phosphorylation pathway and the orchestration of apoptotic cell death [65]. ROS are consistently generated within mitochondria via the electron transport chain. However, enzymatic degradation of neurotransmitters, neuroinflammatory mediators, and redox reactions can also contribute to ROS production [66]. In the anterior chamber, the trabecular meshwork (TM) is the most susceptible tissue to oxidative injury [67]. Oxidative stress on the TM can induce numerous detrimental effects, including altering cell-cycle progression [68,69], changes in the extracellular matrix [70], rearrangement of TM cell cytoskeletal structures, loss of cell-matrix adhesion [71], and alteration of membrane permeability [72]. Additionally, it can induce inflammatory cytokine release and initiate RGC apoptosis [49]. Mitochondrial dysfunction augments the production of ROS while concurrently diminishing the synthesis of ATP. Neuronal cells, which demand substantial energy to sustain electrochemical gradients crucial for signal conduction, are exceedingly susceptible to perturbations in mitochondrial function. The generation of ATP predominantly occurs within the mitochondria via the oxidative phosphorylation mechanism of the electron transport chain, complemented by the glycolytic process [73]. Defects in mitochondrial complex 1 were found to induce ROS release and decrease ATP levels in human glaucomatous TM cells [74]; a study found that RGC death occurred 3–4 months after elevated IOP, with an 18% reduction in mitochondrial membrane potential [75]. Dynamin-related protein 1 (Drp1), which is present on the outer mitochondrial membrane, has been shown to play a role in glaucoma [76]. A recent study showed that the ERK1/2-Drp1-ROS axis can cause mitochondrial dysfunction and apoptosis in RGCs [77]. The involvement of mitochondria in glial neuroinflammation processes is facilitated by the activation of NF-κB through the generation of mitochondrial ROS, resulting in the production of inflammatory cytokines [78].
In response to cellular injury and oxidative stress, numerous pro-inflammatory mediators are upregulated. Normally, the eye is regarded as an immune-privileged site; however, the permeability of the blood–retina barrier may be altered by factors such as oxidative stress, vascular endothelial growth factor, and inflammation [79]. In addition to chemokines and adhesion molecules, cytokines such as interleukins 1β (IL-1β) and tumor necrosis factor-alpha (TNF-α) exhibit increased levels in glaucomatous tissues [80]. Elevated IOP can trigger an innate immune response with researchers finding an increase in microglia activity and cell density as well as expression of the complement of C1q in the retina and optic nerve in mice with glaucoma [81,82]. Adaptive immunity of glaucoma was seen when heat shock protein (HSP)-specific memory T cells were induced by commensal microflora [83]. HSP-27, HSP-60, and HSP-70 autoantibodies have been found in the sera of glaucoma patients [84]. In addition, microglial cells are activated in response to elevated IOP by producing cytokines, mediators, and enzymes that can alter the ECM [85]. Microglial cells can exhibit two phenotypes, M1 (pro-inflammatory) or M2 (neuroprotective); however, in glaucomatous eyes, the majority of microglia do not show the M2 phenotype, and the activities of microglia may have detrimental consequences for the glaucomatous optic disc [86].
In conclusion, the RGC vulnerability hypotheses in glaucoma are multifaceted, encompassing mechanical, vascular, metabolic, excitotoxic, and inflammatory dimensions. The combination of these theories highlights the complexity of glaucomatous neurodegeneration and necessitates a holistic understanding of the changes in RGC functionality, thereby guiding the development of nuanced therapeutic strategies aimed at reducing RGC vulnerability and preserving visual function.
산화 스트레스와 신경 염증은
녹내장의 발병과 관련이 있으며,
활성 산소종(ROS)과 염증 매개체가 세포 손상을 촉진하고
흥분성 환경에 기여하여 RGC 사멸을 악화시킵니다.
녹내장성 눈에서 ROS 생성이 증가하는 데는 여러 가지 요인이 기여할 수 있습니다. 허혈-재관류 손상, 안압 상승, 미토콘드리아 기능 장애는 시신경 머리와 망막에서 ROS의 과잉 생산에 기여하는 주요 요인 중 하나입니다 [49,64]. 미토콘드리아의 주요 역할은 산화적 인산화 경로를 통한 아데노신 삼인산(ATP)의 합성과 세포 사멸의 조율을 포함합니다 [65]. ROS는 전자 전달 사슬을 통해 미토콘드리아 내에서 지속적으로 생성됩니다. 그러나 신경전달물질, 신경염증 매개체의 효소적 분해 및 산화 환원 반응도 ROS 생성에 기여할 수 있습니다 [66]. 전실에서 섬유주망(TM)은 산화적 손상에 가장 취약한 조직입니다 [67]. TM에 대한 산화 스트레스는 세포 주기 진행 변화[68,69], 세포 외 기질 변화[70], TM 세포 골격 구조 재배열, 세포-기질 접착력 상실[71], 막 투과성 변화[72] 등 수많은 해로운 영향을 유발할 수 있습니다. 또한 염증성 사이토카인 방출을 유도하고 RGC 세포 사멸을 시작할 수 있습니다[49]. 미토콘드리아 기능 장애는 ROS의 생성을 증가시키는 동시에 ATP의 합성을 감소시킵니다. 신호 전도에 중요한 전기 화학적 기울기를 유지하기 위해 상당한 에너지를 필요로 하는 신경 세포는 미토콘드리아 기능의 교란에 매우 취약합니다. ATP 생성은 주로 전자 전달 사슬의 산화적 인산화 메커니즘을 통해 미토콘드리아 내에서 일어나며, 해당 과정으로 보완됩니다[73]. 미토콘드리아 복합체 1의 결함은 인간 녹내장 TM 세포에서 ROS 방출을 유도하고 ATP 수준을 감소시키는 것으로 밝혀졌습니다 [74]; 한 연구에 따르면 안압 상승 후 3-4개월이 지나면 미토콘드리아 막 전위가 18% 감소하면서 RGC 사멸이 발생했습니다 [75]. 외부 미토콘드리아 막에 존재하는 다이나민 관련 단백질 1(Drp1)은 녹내장에서 중요한 역할을 하는 것으로 나타났습니다 [76]. 최근 연구에 따르면 ERK1/2-Drp1-ROS 축이 RGC에서 미토콘드리아 기능 장애와 세포 사멸을 일으킬 수 있다고 합니다 [77]. 신경교 신경염증 과정에서 미토콘드리아의 관여는 미토콘드리아 ROS의 생성을 통한 NF-κB의 활성화에 의해 촉진되어 염증성 사이토카인의 생성을 초래합니다 [78].
세포 손상과 산화 스트레스에 대한 반응으로 수많은 전 염증 매개체가 상향 조절됩니다. 일반적으로 눈은 면역력이 약한 부위로 간주되지만 산화 스트레스, 혈관 내피 성장 인자 및 염증과 같은 요인에 의해 혈액-망막 장벽의 투과성이 변경될 수 있습니다 [79]. 케모카인과 접착 분자 외에도 인터루킨 1β(IL-1β) 및 종양괴사인자-알파(TNF-α)와 같은 사이토카인은 녹내장 조직에서 증가된 수치를 보입니다 [80]. 안압 상승은 선천성 면역 반응을 유발할 수 있으며, 연구자들은 녹내장이 있는 마우스에서 망막과 시신경에서 미세아교세포 활성과 세포 밀도가 증가하고 C1q 보체의 발현이 증가하는 것을 발견했습니다 [81,82]. 녹내장의 적응 면역은 공생 미생물에 의해 열충격 단백질(HSP) 특이적 기억 T 세포가 유도되었을 때 나타났습니다 [83]. 녹내장 환자의 혈청에서 HSP-27, HSP-60 및 HSP-70 자가 항체가 발견되었습니다 [84]. 또한 미세아교세포는 안압 상승에 반응하여 사이토카인, 매개체, 효소를 생성하여 ECM을 변화시킬 수 있습니다 [85]. 미세아교세포는 M1(염증성) 또는 M2(신경 보호성)의 두 가지 표현형을 나타낼 수 있지만 녹내장 눈에서는 대부분의 미세아교세포가 M2 표현형을 보이지 않으며 미세아교세포의 활동은 녹내장 시신경에 해로운 결과를 초래할 수 있습니다 [86].
결론적으로 녹내장의 RGC 취약성 가설은 기계적, 혈관적, 대사적, 흥분성 및 염증적 차원을 포괄하는 다면적인 가설입니다. 이러한 이론의 조합은 녹내장성 신경 퇴행의 복잡성을 강조하고 RGC 기능의 변화에 대한 전체적인 이해가 필요하며, 이를 통해 RGC 취약성을 줄이고 시각 기능을 보존하기 위한 미묘한 치료 전략의 개발을 안내합니다.
4. Imaging Technique for Assessing Retinal Ganglion Cells
4.1. Structural Imaging Modalities
The elucidation of RGC structure and function in glaucoma requires cutting-edge imaging techniques that provide precise, dependable, and non-invasive evaluations. OCT is a pioneering modality introduced in the early 1990s, as it allows for the detailed visualization of retinal layers and quantification of the retinal nerve fiber layer (RNFL) [87]. It first produced images through measurement of the echo time delay of light that was backscattered and reflected from ocular tissues [88]. However, due to its suboptimal resolution and slow scan acquisition, visualization of RGCs was difficult. Later, introduction of spectral-domain OCT (SD-OCT), also known as Fourier-domain OCT (FD-OCT), allowed for higher resolution, faster acquisition scan rate, and enhanced reproducibility. Another type of FD-OCT, swept-source OCT (SS-OCT), permits scanning velocities up to 200,000 A-scans/s and employs a laser that rapidly transverses a wide range of frequencies. A photodetector is utilized to capture the interference pattern instead of the spectrometer used in SD-OCT [89]. The capability of OCT to detect subtle alterations in RNFL thickness provides invaluable insights into early glaucomatous changes and is a crucial tool for monitoring disease progression and treatment response [90].
Moreover, confocal scanning laser ophthalmoscopy (cSLO) is widely used to evaluate the optic nerve head topography, producing detailed images of the optic disc and its adjacent tissues that are crucial for assessing structural changes indicative of glaucomatous damage [91]. It consists of a 670 nm diode laser, which uses a narrow laser beam and small aperture to limit scattered light from outside the focal plane [2]. Unlike standard fundus imaging, this provides excellent lateral resolution for topography images but poor axial resolution.
Adaptive optics (AO) compensates for wavefront distortions with a deformable mirror and reduces the optical aberrations that can cause blurring and image artifacts in ophthalmic imaging [92]. In 1997, AO was integrated with a fundus camera, with scanning laser ophthalmoscopy (SLO) in 2002, and with OCT in 2005. Pairing AO with OCT enables three-dimensional, diffraction-limited images of the optic nerve head, lamina cribrosa, and retina. It is noteworthy that individual nerve fiber bundles can be distinguished in these images, even after repeated sessions, which renders this method appealing for investigating the progression of glaucoma [93].
4.2. Functional Imaging Modalities
Functional evaluation of RGCs is of equal importance, and developments in electroretinography (ERG) have enabled the isolated evaluation of RGC function, providing a unified view of both structural and functional integrity. Recent findings have shown that pattern reversal ERG (PERG), photopic negative response (PhNR), and multifocal ERG can be objective measures of RGCs [94]. At present, PERG is one of the most recognized ERG methods for the identification of glaucoma. Earlier investigations have suggested that it could be particularly advantageous when used in conjunction with the diagnosis and treatment of individuals suspected of having glaucoma, particularly those with normal or borderline visual fields and/or RNFL thickness [95,96]. A study found that it takes 2 years for PERG to change by 10% and 10 years for RNFL to change by 10%, indicating a time lag of 8 years between changes in PERG and RNFL in glaucoma suspects [95]. Another study identified individuals with glaucoma four years prior to the onset of visual field alterations with a sensitivity and specificity of 75% and 76%, respectively [96]. Furthermore, the prognostic capability of PERG in the evaluation of glaucomatous changes has been observed [97,98]. Research indicated that lower pERG N95 amplitudes correlate with retinal nerve fiber layer defects in preperimetric glaucoma and decrease progressively from normal controls to perimetric glaucoma patients, suggesting that pERG N95 amplitude could be a useful metric for early glaucoma detection [97].
PhNR is a slow negative wave after the b-wave in ERG under photopic conditions that originates from RGCs [99]. The amplitude of PhNR and the PhNR/b-wave ratio decreases with the advancement of glaucoma, as assessed by visual field defect increases [100]. Multifocal ERG, introduced in 1992, enables numerous localized retinal responses to be identified from a single recording [101]. Research findings indicate that individuals diagnosed with glaucoma or with suspected glaucoma may exhibit diminished amplitude and/or delayed implicit time in their mfERG responses, particularly in regions that align with visual field impairments [102,103]. Several investigations have employed mfERG to differentiate between glaucoma and healthy eyes. However, thus far, none of this research has provided conclusive evidence about the diagnostic utility of the mfERG approach in glaucoma [101,104,105].
In addition, Standard Automated Perimetry (SAP) is widely regarded as a gold standard in the diagnosis, monitoring, and management of glaucoma. The field of perimetry progressed in the 1930s, when Goldmann introduced standardized kinetic perimetry. The stimuli were projected onto a hemispheric bowl and systematically moved from areas where they were not perceived to locations where they were initially detected. However, kinetic perimetry is subjective, relying on training and experience. There are significant technological restrictions, including psychophysical sensitivity to moving targets, spatial summation effects, and subject response time to the examiner. SAP, an example of static perimetry, uses a contrast stimulus. RGCs increase their firing rate when the stimulus contrast increases, until the observer detects this and responds [106]. SAP is not selective for a specific RGC type; there are newer tests that demonstrate and target specific RGCs, such as short-wavelength automated perimetry (SWAP) and frequency doubling technology (FDT). SWAP differs from SAP in its blue-light stimulus on yellow background illumination and assesses the short-wavelength cone pathway. These blue-yellow ganglion cells extend their axons towards the interlaminar, koniocellular layers of the LGN [107,108]. Researchers suggest SWAP can detect functional damage in glaucoma earlier than SAP [109,110]. Ocular hypertension patients at higher risk for glaucoma have more SWAP abnormalities than those at lesser risk [111]. However, SWAP tests take longer, which reduces patient satisfaction and healthcare practice efficiency. It may take 15% longer duration for a full-threshold SWAP than a full-threshold SAP. The introduction of the SITA threshold technique has overcome this limitation and significantly reduced test length. However, SWAP can be affected by media opacities and has greater intra-test and inter-test variability [112]. FDT measures contrast sensitivity for frequency-doubling stimulus, which is assumed to be mediated by a subpopulation of magnocellular RGCs. Several studies have demonstrated that FDT has high sensitivity and specificity in distinguishing glaucomatous patients from normal subjects, and test outcomes can be used to predict location and future onset of functional loss in suspected glaucoma as determined by SAP [113,114]. When compared to SAP and SWAP, FDT has a shorter testing time and lower variability, which may be attributed to the smaller influence of media opacities, pupil diameter, and refractive errors during testing [115].
A further advanced imaging modality utilized in the field of glaucoma is the Detection of Apoptosing Retinal Cells (DARC), now undergoing phase two clinical trials [116]. The method involves the intravenous administration of a fluorescent marker called Annexin V, which exhibits a strong binding affinity for phosphatidylserine. This molecule is expressed on the cell surface in apoptosis. The marker is taken up by retinal cells, which can be observed on cSLO, enabling the identification of retinal ganglion cells that are undergoing apoptosis [116]. In vivo studies have shown compatible DARC labeled cells correlated with RGC apoptosis [117,118].
Table 2 delineates the structural and functional imaging modalities employed for the assessment of RGCs in glaucoma. The refinement and integration of these techniques are indispensable for improving the diagnostic accuracy and elucidating the complex pathophysiology of RGCs in glaucomatous conditions, thereby molding the therapeutic intervention and management strategies.
Table 2
Imaging modalities for assessing structural and functional retinal ganglion cells in glaucoma.
Retinal ImagingDescriptionApplicationsAdvantagesDisadvantages
| Optical coherence tomography (OCT) | A non-invasive imaging technique that uses light waves to take high-resolution cross-sectional pictures of the retina. | Measurement of RNFL 1 thickness, optic nerve, and macular ganglion cell complex | High-resolution images and non-invasive | Affected by media opacities (e.g., cataract), poor visualization of RGCs 2 |
| Confocal scanning laser ophthalmoscopy (cSLO) | cSLO utilizes a narrow laser beam to scan the retina point by point through a small aperture. | Visualization of optic nerve head | High contrast image, excellent lateral resolution, and monitoring of RNFL defects and glaucomatous changes | Affected by media opacities (e.g., cataract) and poor axial resolution |
| Adaptive optics imaging | Adaptive optics imaging employs deformable mirrors and wavefront sensors to correct optical aberrations. | Visualization of RGC | High-resolution, allowing visualization of individual cells, and can correct optical aberrations | Pupil dilation, longer time needed, and costly |
| Electroretinography (ERG) | ERG measures the electrical responses of various cell types in the retina after visual stimulation. | Assesses the functional status of RGCs using PERG 3, PhNR 4, and mfERG 5 | Functional assessment of RGCs, objective measurement of retinal activity, may detect early glaucomatous changes | Affected by media opacities, interpretation based on test parameters, utilization still under evaluation |
| Standard automated perimetry (SAP) | SAP assesses the visual field by presenting light stimuli of varying intensities in a standardized pattern to detect visual deficits. | The central tool in glaucoma diagnosis, staging, and monitoring. Visual field defects correlate with RGC loss and damage to the optic nerve. | Widely accepted and utilized, objective measures of visual function, able to monitor progression, offering other specific techniques in assessing specific RGC types, such as SWAP 6 and FDT 7 | Dependent on patient’s cooperation and understanding, can be affected by media opacities and ptosis, test–retest variability |
| Detection of apoptosing retinal cells (DARC) | DARC utilizes fluorescently tagged annexin-V intravenously to mark apoptotic RGCs, which are then identified with cSLO. | To assess efficacy of therapeutic agents | Identification of individual apoptosis RGC cells and early detection of glaucomatous damage | Invasive, long-term effects not yet known |
1 RNFL, retinal nerve fiber layer; 2 RGC, retinal ganglion cell; 3 PERG, pattern reversal ERG; 4 PhNR, photopic negative response; 5 mfERG, multifocal ERG; 6 SWAP, short-wavelength automated perimetry; 7 FDT, frequency doubling technology.
5. Therapeutic Approaches Targeting Retinal Ganglion Cells
Therapeutic efforts concentrating on RGCs are essential for addressing the multifaceted nature of glaucoma, with the goal of slowing the disease’s progression by emphasizing cellular protection, restoration, and resilience. Although IOP reduction is the only treatment currently proven to be clinically effective in managing glaucoma, it is considered indirectly neuroprotective. Nonetheless, the development of direct neuroprotective therapies targets the neurons, specifically the RGCs themselves. A range of therapeutic approaches designed to preserve and restore RGCs have been explored within the research context for glaucoma treatment (Table 3).
Table 3
Therapeutic targets for retinal ganglion cells.
CategoryAgent/MethodEffect/OutcomeReference
| Neuroprotective agents | Brimonidine | Alpha-2 adrenergic receptor agonist; reduces extracellular glutamate, blocks NMDA 1 receptor, and supports RGC 2 survival under high IOP 3 | [119,120,121,122,123,124] |
| MK801 (dizocilpine maleate) | Strong glutamate inhibitor; uncompetitive NMDA antagonist with neuroprotective effects; can be neurotoxic due to long half-life | [125,126] | |
| Memantine | Selective and non-competitive NMDA receptor inhibitor; showed partial RGC protection in preclinical studies but no significant benefit in Phase 3 clinical trial | [127,128,129] | |
| Calcium channel blockers | Topical 2% flunarizine | Reduced RGC injury in rabbits under high IOP conditions | [130] |
| Brovincamine and nilvadipine | Showed improved visual field progression and increased posterior choroidal circulation | [131,132] | |
| Anti-oxidative agents | Coenzyme Q10 (CoQ10) | Protects RGCs from oxidative stress, supports ATP 4 synthesis, inhibits ROS production; promotes RGC survival in oxidative stress models | [45,133,134,135,136,137] |
| Neurotrophic factors | Brain-derived neurotrophic factor (BDNF) | Intravitreal injection protects RGCs in animal models; restores PERG and VEP damage; increases RGC survival in hypertensive rat eyes with intraocular injections | [138,139,140,141,142] |
| Ciliary neurotrophic factor (CNTF) | Expressed in Muller cells; offers protection against NO-induced 5 cell death and optic nerve axotomy in rats | [143,144,145,146] | |
| Nerve growth factor (NGF) | NGF eye drops can mitigate glaucoma-associated optic nerve damage in rats, with higher RGC survival; topical rh-NGF 6 results in improved RGC survival, decreased apoptosis, and reduced astrocyte activity in the optic nerve in a rat model | [147,148,149] | |
| Glial cell line–derived neurotrophic factor (GDNF) | Enhances GLAST gene expression in Müller glia, essential for RGC protection; GDNF microspheres increase RGC survival and density in rat models | [150,151,152] | |
| Gene therapy | γ-synuclein (mSncg) promoter | Preserves the acutely injured RGC somata and axons | [153] |
| Complement C3 | Neuroprotection of RGC axons and somata | [154] | |
| Calcium/calmodulin-stimulated protein kinase II (CaMKII) | Protection of RGCs and their axons | [155] | |
| Nicotinamide mononucleotide adenylyl transferase (NMNAT) | Significant neuroprotection of both RGC soma and axon and preservation of visual function | [156] | |
| X-linked inhibitor of apoptosis (XIAP) | Provide both functional and structural protection of RGC | [157] | |
| BCLXL | Robustly attenuate both RGC soma pathology and axonal degeneration in the optic nerve | [158] | |
| Stem cell Therapy | Brain-derived, hippocampal-derived stem cells | Differentiate into amacrine and horizontal cells | [159] |
| Muller glia stem cells | Differentiate into RGCs, restore connectivity to other neurons, and address negative scotopic threshold response | [160] | |
| IOP management | Trabeculectomy and Pharmacological Medications | Essential for reducing mechanical stress on RGCs, preventing further degeneration |
1 NMDA, N-methyl-d-aspartate; 2 RGC, retinal ganglion cell; 3 IOP, intraocular pressure; 4 ATP, adenosine triphosphate; 5 NO, nitric oxide; 6 rh-NGF, recombinant human nerve growth factor.
5.1. Neuroprotective Agents
Neuroprotective agents such as brimonidine and memantine have demonstrated promise in preventing RGC degeneration by modulating neurotransmitter activity, reducing excitotoxicity, and mitigating oxidative stress. The excitotoxicity theory mentioned previously contributes to RGC degeneration through excessive glutamate release and overactivation of the NMDA receptor, and many studies have suggested the possibility of blocking the glutamate receptor as a method for RGC neuroprotection. Alpha-adrenergic receptors have been identified within RGCs, and immunohistochemical analyses demonstrated the presence of alpha-adrenergic receptors in human, bovine, and porcine retinas as well as the inner nuclear layer of the rat retina [161,162]. Brimonidine, an alpha-2 adrenergic receptor agonist, can increase retinal metabolism and neuronal growth in cultured retinal cells as well as diminish extracellular glutamate and NMDA receptor blockage [119,120,121]. A study found that continuous subcutaneous brimonidine treatment increased RGC survival under high IOP pressure for 8 weeks [122]. In addition, the Low-Pressure Glaucoma Treatment Study (LoGTS), which randomized patients to either topical brimonidine or timolol, showed 9% visual field progression in the brimonidine group and 39% progression in the timolol group [123]. However, this study had significantly greater numbers of drop-outs in the brimonidine group. In another study, brimonidine 0.2% caused less RNFL loss than timolol 0.5% in ocular hypertension patients during a 12-month period [124], which suggests the neuroprotective effect of brimonidine. MK801 (dizocilpine maleate), a strong glutamate inhibitor, is an uncompetitive NMDA antagonist with neuroprotective effects for RGC; nevertheless, due to its lengthy half-life, it can be neurotoxic [125,126]. Memantine, on the other hand, selectively and non-competitively inhibits the NMDA receptor with moderate affinity. In preclinical and experimental studies, memantine showed a partial protective effect of RGCs against glutamate toxicity when administered with low-dose glutamate in rats and enhanced RGC survival with high IOP in monkeys [127,128]. However, a Phase 3 clinical trial did not find any significant benefits compared with the placebo [129].
5.2. Calcium Channel Blockers
Calcium channel blockers (CCBs) have been examined for their neuroprotective effects against RGCs. Calcium influx into neural cells can cause apoptosis and cell death when NMDA is activated; hence, CCB has been studied for neuroprotection in glaucoma. Topical 2% flunarizine, a calcium channel blocker, reduced rabbit retinal RGC injury under high IOP ischemia conditions [130]. Brovincamine and nilvadipine have high blood–brain barrier permeability and have been shown to improve visual field progression and increase posterior choroidal circulation [131,132]. Although CCB appears promising, additional research is needed to determine its therapeutic effectiveness, as it may disrupt the autoregulation of blood flow during an acute spike in IOP [163].
5.3. Antioxidants
Oxidative stress with the increase of ROS can be detrimental to RGCs. Coenzyme Q10 (CoQ10), an electron transport chain cofactor, stabilizes mitochondrial membrane potential, supports adenosine triphosphate (ATP) synthesis, and inhibits ROS production to protect neuronal cells from oxidative stress [133,134]. CoQ10 has been shown to protect RGCs during hydrogen peroxide oxidative stress in vitro and NMDA-induced glutamate excitotoxicity in vivo [135]. It can also promote RGC survival by 29% in mouse models under oxidative stress [136]. A study found that topical Coqun eye drops (CoQ10 and vitamin E) in glaucoma patients demonstrated improved inner retinal function, indicated by PERG and visual cortex responses as determined by visual evoked potential (VEPs) [137].
5.4. Neurotrophic Factors
Beyond pharmacologic strategies, neurotrophic factor supplementation has attracted considerable interest, with a focus on the potential of BDNF, CNTF, NGF, and GDNF to promote RGC survival and resilience. These factors have demonstrated efficacy in enhancing RGC survival, reducing apoptosis, and promoting cellular resilience, highlighting their therapeutic potential in the treatment of glaucoma. Studies have shown that intravitreal BDNF injection protects RGCs in rat and primate optic nerve injury models [138,139]. In chronic intraocular hypertension, recombinant human BDNF eye drops restored PERG and VEP damage [140]. Intraocular injections of BDNF at 1.0 g/L in moderately chronic hypertensive rat eyes increased RGC survival by 2 weeks [141]; however, high-dose BDNF may significant downregulate TrkB expression, therefore lowering BDNF efficiency [142]. CNTF is particularly expressed in Muller cells, and its neuroprotective effect is mediated by glial cells, which respond by producing neurotrophic factors such as basic fibroblast growth factor [143,144]. A single vitreous injection of CNTF protein protected RGCs from nitric oxide (NO)-induced cell death and optic nerve axotomy in rats [145,146].
Prior research has indicated a correlation between RGC depletion and the decreased expression of NGF and its receptor in the retina. Ocular administration of NGF notably mitigated the impairments instigated by glaucoma [147]. Lambiase et al. demonstrated that NGF eye drops can mitigate glaucoma-associated optic nerve damage in rats, with higher RGC survival in treated groups with the administration of NGF eye drops four times a day over a span of 7 weeks [148]. In a separate investigation, recombinant human nerve growth factor (rh-NGF) was employed in a rat model. When rh-NGF was applied topically twice a day for 3 weeks, there was a marked enhancement in RGC survival, evidenced by diminished RGC apoptosis and an increased RGC count in the inferior retina. Additionally, topical rh-NGF was found to bolster axonal survival and curtail astrocyte activity within the optic nerve [149].
In Müller glia, GDNF intriguingly augments GLAST gene expression, which is crucial for RGC safeguarding. In mice with GLAST and EAAC1 knockouts, RGC mortality and glaucomatous injury emerged, even without elevated ocular pressure [164]. Employing this strategy, post-microsphere injections laden with GDNF, RGC longevity in DBA/2J mice was extended for nine months [150]. GDNF/Vitamin E Poly(lactide-co-glycolide) (PLGA) microsphere injections in a glaucoma animal model notably improved RGC longevity over 11 weeks, surpassing GDNF, Vitamin E, and mere microspheres [151]. Ward et al. established that GDNF via microspheres notably amplified RGC persistence in a spontaneous glaucoma model, showcasing a 3.5-fold RGC density increment in 15-month survival compared to non-treated counterparts [152]. BDNF-mediated, CNTF-mediated, NGF-mediated, and GDNF-mediated neuroprotection in glaucoma has great potential, but retinal delivery is challenging, and adverse effects must be addressed.
5.5. Gene Therapy
The retina presents an optimal site for gene interventions due to its accessibility, distinct functional indicators, semi-immune protection, and localized non-systemic position [165]. Recent studies suggest the potential of gene therapy in preserving both the architecture and activity of RGC.
The γ-Synuclein (mSncg) promoter is a notable target and is of great relevance to RGCs. Through a combination of adeno-associated virus (AAV)-mSncg promoter and clustered, regularly interspaced short palindromic repeats (CRISPR)/Cas9 gene editing, pro-degenerative genes can be knocked down. Wang et al., using an AAV2-mSncg in human pluripotent stem cell (hPSC)-derived RGCs and a mice optic nerve crush model, effectively preserved the acutely injured RGC somata and axons [153].
Complement C3 is another target in gene therapy focused on RGC preservation. By overexpressing the C3 inhibitor, the activation of complement C3d is reduced. Research by Bosco et al. [154] combined AAV retinal gene therapy with the targeted C3 inhibitor through intravitreal injections in a mice glaucoma model. The result revealed neuroprotection of RGC axons and somata despite continued intraocular pressure elevation.
In relation to RGC health, CaMKII (Calcium–calmodulin ((CaM))-dependent protein kinase II) assumes a crucial function. Augmenting the expression of CaMKII provides protection to both RGCs and their associated axons. This protective mechanism is realized through an AAV-mediated treatment involving CaMKIIa Threonine 286D, administered via intravitreal injection in a mouse glaucoma model [155]. Subsequent findings indicate that the reactivation of CaMKII safeguards the extensive axonal projections of RGCs in vivo, maintaining both visual functionalities, spanning from the retina to the visual cortex, and the integrity of visually driven behavior.
NMNAT (nicotinamide mononucleotide adenylyl transferase) represents another gene target critically linked to the well-being of RGCs. As illustrated by Fang et al., the overexpression of the long half-life NMNAT2 mutant, steered by the RGC-specific mSncg promoter in mice, facilitated by AAV2 intravitreal injection, reinstates the diminished nicotinamide adenine dinucleotide (NAD+) levels in RGCs and ONs affected by glaucoma. This RGC-centric gene therapy approach confers notable neuroprotective benefits to the RGC soma and its axon, while ensuring the maintenance of visual functionality in the context of the optic nerve crush model [156].
The X-linked inhibitor of apoptosis (XIAP) presents an alternative pathway for safeguarding RGCs. Through inhibition of apoptotic activation, RGCs gain both structural and functional defense. This method, implemented through intravitreal injection in a murine glaucoma paradigm, was investigated by Visuvanathan et al. [157]. Subsequent findings from PERGs demonstrated that XIAP treatment substantially preserved the function of somal and axonal RGCs in glaucomatous eyes.
The BCLXL gene emerges as a potential therapeutic target for RGCs. The activation of the BAX protein is pivotal in the intrinsic apoptotic pathway leading to RGC demise in glaucoma, with BCLXL serving as its primary counteragent. Donahue et al. engineered a mCherry-BCLXL fusion protein, inhibiting BAX localization and activation in mitochondrial tissue culture cells. Upon packaging into AAV2 and administering intravitreally in a murine glaucoma model, there was no alteration in the IOP trajectory compared to control mice. However, notable reductions were observed in RGC soma pathology and axonal deterioration in the optic nerve [158].
5.6. Stem Cell Therapy
Emerging evidence suggests the relevance of stem cell therapy for RGC regeneration. The potential of therapy derived from stem cells resides in its capacity to replace damaged RGCs and restore retinal functionality; however, this would require the cells to differentiate into RGCs and become integrated and capable of synapsing in precise brain locations. Stem cell transplantation from brain-derived, hippocampal-derived, and bone marrow–derived cells mostly differentiates into amacrine and horizontal cells [159]. However, a study found that Muller glia can be differentiated into RGCs, and they were able to restore the connectivity to other neurons and the negative scotopic threshold response [160]. The road to clinical application is fraught with difficulties, including assuring transplanted cells’ long-term safety and stability, optimizing delivery systems, and achieving functional integration with existing brain networks.
5.7. Intraocular Pressure Management
Management of IOP remains the foundational strategy in the treatment of glaucoma, with medical therapy typically serving as the initial approach to decrease IOP. Pharmacological agents, often in the form of topical eye drops, are the first line of defense to mitigate the mechanical stress exerted on RGCs and avert further deterioration and loss. Should medication not suffice, laser procedures are considered the subsequent step in the therapeutic cascade. If these measures prove inadequate, surgical interventions such as trabeculectomy or minimally invasive glaucoma surgeries (MIGS) are employed to further lower IOP. In cases where conventional surgeries do not achieve the desired outcomes, the implantation of shunts or valves may be warranted as a final recourse to manage the disease effectively [166].
5.8. Blood Flow Enhancement
Regeneration of the optic nerve may be facilitated by enhancing blood flow and providing robust vascular support. Optimal blood circulation is crucial for delivering nutrients and oxygen, which are essential for the survival and repair of neurons. When the optic nerve is damaged, improving blood flow can mitigate nerve degeneration and facilitate the transport of neurotrophic factors and other regenerative molecules to the site of injury. Vascular support, particularly through the stabilization of blood vessels and the prevention of ischemic damage, can further potentiate the regeneration of optic nerve fibers. Therapeutic approaches, though still largely investigational, are aimed at augmenting these physiological processes to aid in repair and regeneration. For example, ginkgo biloba extract (GBE) has been explored for its potential to improve ocular blood flow [167]. EGb 761, a refined extract of ginkgo biloba, is characterized by a reduction in ginkgolic acids, which are considered undesirable, while retaining significant concentrations of bioactive constituents, specifically flavonoid glycosides (comprising 22.0–27.0%) and terpene lactones (encompassing 5.0–7.0%) [168]. These flavonoid compounds, known for their antioxidant properties, play a role in maintaining vascular integrity, potentially offering advantages for the perfusion of the optic nerve [169]. Research involving a murine model with sustained moderate IOP elevation revealed that, following a 5-month regimen of EGb 761 administration, there was no observable impact on IOP; however, there was a notable attenuation in the degeneration of RGCs [168]. Complementary interventions, including lifestyle modifications that encourage exercise and a balanced diet, support overall vascular integrity. While these modalities offer hope, their integration into clinical practice necessitates further validation through rigorous research to establish efficacy and safety.
In conclusion, the combination of diverse therapeutic strategies, including pharmacological, neurotrophic, stem cell, and gene therapy approaches, broadens the spectrum of interventions available to clinicians, thereby facilitating the development of individualized treatment plans based on the needs and disease characteristics of each patient. These evolving therapies highlight the need for a multidimensional approach incorporating diverse therapeutic modalities to address the complexities of RGC vulnerability and pave the way for glaucoma management on a global scale.
6. Conclusions and Future Directions
The pursuit of knowledge about RGCs’ role in glaucoma continues to uncover new pathways for research and innovation, necessitating a forward-thinking strategy in both the research and therapeutic arenas. Gaining a comprehensive understanding of the underlying theories pertaining to the pathophysiology of glaucoma elucidates the intricate nature of glaucomatous degeneration and holds significant relevance in identifying potential therapeutic targets for future interventions. The assessment of RGC functionality by several imaging modalities is of utmost importance in the clinical management of glaucoma. The growing interest in the cellular and molecular mechanisms behind RGC degeneration supports the development of novel therapeutic targets and therapies, which may have implications for personalized medicine methods in the management of glaucoma.
Expanding on this, the future of glaucoma research could pivot towards pioneering treatments that accomplish more than halting the progression of the disease—aiming instead to reverse its course. The exploration of neuroregenerative therapies, such as cell-based therapies, offers the tantalizing prospect of not only preserving but also restoring lost vision [170]. Moreover, advancements in nanotechnology hold the potential for targeted drug delivery systems that could administer therapeutic agents directly to RGCs, enhancing efficacy and reducing systemic side effects [171]. Artificial intelligence and machine learning algorithms could also revolutionize early detection and personalized treatment plans based on predictive modeling [172]. As we stand on the cusp of these scientific horizons, it is paramount that our research strategies incorporate these emerging modalities, ensuring a robust and dynamic approach to overcoming the challenges posed by glaucoma.
Funding Statement
This research received no external funding.
Author Contributions
Conceptualization, K.M.F. and D.-W.L.; validation, Y.-H.C. and D.-W.L.; resources, D.-W.L.; writing—original draft preparation, K.M.F. and T.-H.T.; writing—review and editing, Y.-H.C. and D.-W.L.; visualization, K.M.F. and T.-H.T.; supervision, Y.-H.C. and D.-W.L.; funding acquisition, D.-W.L. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Ethical review and approval were waived for this study due to due to the nature of the article as a review article.
Informed Consent Statement
Patient consent was waived due to the nature of the article as a review article.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Footnotes
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content.
References
1. Tham Y.C., Li X., Wong T.Y., Quigley H.A., Aung T., Cheng C.Y. Global prevalence of glaucoma and projections of glaucoma burden through 2040: A systematic review and meta-analysis. Ophthalmology. 2014;121:2081–2090. doi: 10.1016/j.ophtha.2014.05.013. [PubMed] [CrossRef] [Google Scholar]
2. Balendra S.I., Normando E.M., Bloom P.A., Cordeiro M.F. Advances in retinal ganglion cell imaging. Eye. 2015;29:1260–1269. doi: 10.1038/eye.2015.154. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
3. Dias M.S., Luo X., Ribas V.T., Petrs-Silva H., Koch J.C. The Role of Axonal Transport in Glaucoma. Int. J. Mol. Sci. 2022;23:3935. doi: 10.3390/ijms23073935. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
4. Dunaief J.L., Dentchev T., Ying G.S., Milam A.H. The role of apoptosis in age-related macular degeneration. Arch. Ophthalmol. 2002;120:1435–1442. doi: 10.1001/archopht.120.11.1435. [PubMed] [CrossRef] [Google Scholar]
5. Boia R., Ruzafa N., Aires I.D., Pereiro X., Ambrosio A.F., Vecino E., Santiago A.R. Neuroprotective Strategies for Retinal Ganglion Cell Degeneration: Current Status and Challenges Ahead. Int. J. Mol. Sci. 2020;21:2262. doi: 10.3390/ijms21072262. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
6. Vernazza S., Oddone F., Tirendi S., Bassi A.M. Risk Factors for Retinal Ganglion Cell Distress in Glaucoma and Neuroprotective Potential Intervention. Int. J. Mol. Sci. 2021;22:7994. doi: 10.3390/ijms22157994. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
7. Masland R.H. The neuronal organization of the retina. Neuron. 2012;76:266–280. doi: 10.1016/j.neuron.2012.10.002. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
8. Martin P.R. Colour processing in the primate retina: Recent progress. Pt 3J. Physiol. 1998;513:631–638. doi: 10.1111/j.1469-7793.1998.631ba.x. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
9. Manookin M.B., Patterson S.S., Linehan C.M. Neural Mechanisms Mediating Motion Sensitivity in Parasol Ganglion Cells of the Primate Retina. Neuron. 2018;97:1327–1340.e1324. doi: 10.1016/j.neuron.2018.02.006. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
10. Weber A.J., Kaufman P.L., Hubbard W.C. Morphology of single ganglion cells in the glaucomatous primate retina. Invest. Ophthalmol. Vis. Sci. 1998;39:2304–2320. [PubMed] [Google Scholar]
11. Martin P.R., Grunert U. Analysis of the short wavelength-sensitive (“blue”) cone mosaic in the primate retina: Comparison of New World and Old World monkeys. J. Comp. Neurol. 1999;406:1–14. doi: 10.1002/(SICI)1096-9861(19990329)406:1<1::AID-CNE1>3.0.CO;2-1. [PubMed] [CrossRef] [Google Scholar]
12. Crook J.D., Peterson B.B., Packer O.S., Robinson F.R., Gamlin P.D., Troy J.B., Dacey D.M. The smooth monostratified ganglion cell: Evidence for spatial diversity in the Y-cell pathway to the lateral geniculate nucleus and superior colliculus in the macaque monkey. J. Neurosci. 2008;28:12654–12671. doi: 10.1523/JNEUROSCI.2986-08.2008. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
13. Do M.T.H. Melanopsin and the Intrinsically Photosensitive Retinal Ganglion Cells: Biophysics to Behavior. Neuron. 2019;104:205–226. doi: 10.1016/j.neuron.2019.07.016. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
14. Sanes J.R., Masland R.H. The types of retinal ganglion cells: Current status and implications for neuronal classification. Annu. Rev. Neurosci. 2015;38:221–246. doi: 10.1146/annurev-neuro-071714-034120. [PubMed] [CrossRef] [Google Scholar]
15. Silveira L.C., Saito C.A., Lee B.B., Kremers J., da Silva Filho M., Kilavik B.E., Yamada E.S., Perry V.H. Morphology and physiology of primate M- and P-cells. Prog. Brain Res. 2004;144:21–46. doi: 10.1016/S0079-6123(03)14402-0. [PubMed] [CrossRef] [Google Scholar]
16. Paknahad J., Loizos K., Yue L., Humayun M.S., Lazzi G. Color and cellular selectivity of retinal ganglion cell subtypes through frequency modulation of electrical stimulation. Sci. Rep. 2021;11:5177. doi: 10.1038/s41598-021-84437-w. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
17. Flammer J., Mozaffarieh M. What is the present pathogenetic concept of glaucomatous optic neuropathy? Surv. Ophthalmol. 2007;52((Suppl. 2)):S162–S173. doi: 10.1016/j.survophthal.2007.08.012. [PubMed] [CrossRef] [Google Scholar]
18. Weinreb R.N., Aung T., Medeiros F.A. The pathophysiology and treatment of glaucoma: A review. JAMA. 2014;311:1901–1911. doi: 10.1001/jama.2014.3192. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
19. Yan D.B., Coloma F.M., Metheetrairut A., Trope G.E., Heathcote J.G., Ethier C.R. Deformation of the lamina cribrosa by elevated intraocular pressure. Br. J. Ophthalmol. 1994;78:643–648. doi: 10.1136/bjo.78.8.643. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
20. Downs J.C., Roberts M.D., Burgoyne C.F. Mechanical environment of the optic nerve head in glaucoma. Optom. Vis. Sci. 2008;85:425–435. doi: 10.1097/OPX.0b013e31817841cb. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
21. Burgoyne C.F., Downs J.C., Bellezza A.J., Suh J.K., Hart R.T. The optic nerve head as a biomechanical structure: A new paradigm for understanding the role of IOP-related stress and strain in the pathophysiology of glaucomatous optic nerve head damage. Prog. Retin. Eye Res. 2005;24:39–73. doi: 10.1016/j.preteyeres.2004.06.001. [PubMed] [CrossRef] [Google Scholar]
22. Shiga Y., Kunikata H., Aizawa N., Kiyota N., Maiya Y., Yokoyama Y., Omodaka K., Takahashi H., Yasui T., Kato K., et al. Optic Nerve Head Blood Flow, as Measured by Laser Speckle Flowgraphy, Is Significantly Reduced in Preperimetric Glaucoma. Curr. Eye Res. 2016;41:1447–1453. doi: 10.3109/02713683.2015.1127974. [PubMed] [CrossRef] [Google Scholar]
23. Caprioli J., Coleman A.L., Blood Flow in Glaucoma D. Blood pressure, perfusion pressure, and glaucoma. Am. J. Ophthalmol. 2010;149:704–712. doi: 10.1016/j.ajo.2010.01.018. [PubMed] [CrossRef] [Google Scholar]
24. Leske M.C., Wu S.Y., Hennis A., Honkanen R., Nemesure B., Group B.E.S. Risk factors for incident open-angle glaucoma: The Barbados Eye Studies. Ophthalmology. 2008;115:85–93. doi: 10.1016/j.ophtha.2007.03.017. [PubMed] [CrossRef] [Google Scholar]
25. Memarzadeh F., Ying-Lai M., Chung J., Azen S.P., Varma R., Los Angeles Latino Eye Study G. Blood pressure, perfusion pressure, and open-angle glaucoma: The Los Angeles Latino Eye Study. Invest. Ophthalmol. Vis. Sci. 2010;51:2872–2877. doi: 10.1167/iovs.08-2956. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
26. Zheng Y., Wong T.Y., Mitchell P., Friedman D.S., He M., Aung T. Distribution of ocular perfusion pressure and its relationship with open-angle glaucoma: The singapore malay eye study. Invest. Ophthalmol. Vis. Sci. 2010;51:3399–3404. doi: 10.1167/iovs.09-4867. [PubMed] [CrossRef] [Google Scholar]
27. Geijer C., Bill A. Effects of raised intraocular pressure on retinal, prelaminar, laminar, and retrolaminar optic nerve blood flow in monkeys. Invest. Ophthalmol. Vis. Sci. 1979;18:1030–1042. [PubMed] [Google Scholar]
28. Alm A., Bill A. Ocular and optic nerve blood flow at normal and increased intraocular pressures in monkeys (Macaca irus): A study with radioactively labelled microspheres including flow determinations in brain and some other tissues. Exp. Eye Res. 1973;15:15–29. doi: 10.1016/0014-4835(73)90185-1. [PubMed] [CrossRef] [Google Scholar]
29. Grieshaber M.C., Mozaffarieh M., Flammer J. What is the link between vascular dysregulation and glaucoma? Surv. Ophthalmol. 2007;52((Suppl. 2)):S144–S154. doi: 10.1016/j.survophthal.2007.08.010. [PubMed] [CrossRef] [Google Scholar]
30. Galambos P., Vafiadis J., Vilchez S.E., Wagenfeld L., Matthiessen E.T., Richard G., Klemm M., Zeitz O. Compromised autoregulatory control of ocular hemodynamics in glaucoma patients after postural change. Ophthalmology. 2006;113:1832–1836. doi: 10.1016/j.ophtha.2006.05.030. [PubMed] [CrossRef] [Google Scholar]
31. Wang X., Wang M., Liu H., Mercieca K., Prinz J., Feng Y., Prokosch V. The Association between Vascular Abnormalities and Glaucoma-What Comes First? Int. J. Mol. Sci. 2023;24:13211. doi: 10.3390/ijms241713211. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
32. Wang Q., Qu X., Chen W., Wang H., Huang C., Li T., Wang N., Xian J. Altered coupling of cerebral blood flow and functional connectivity strength in visual and higher order cognitive cortices in primary open angle glaucoma. J. Cereb. Blood Flow. Metab. 2021;41:901–913. doi: 10.1177/0271678X20935274. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
33. Ghanem A.A., Elewa A.M., Arafa L.F. Endothelin-1 and nitric oxide levels in patients with glaucoma. Ophthalmic Res. 2011;46:98–102. doi: 10.1159/000323584. [PubMed] [CrossRef] [Google Scholar]
34. Shoshani Y.Z., Harris A., Shoja M.M., Rusia D., Siesky B., Arieli Y., Wirostko B. Endothelin and its suspected role in the pathogenesis and possible treatment of glaucoma. Curr. Eye Res. 2012;37:1–11. doi: 10.3109/02713683.2011.622849. [PubMed] [CrossRef] [Google Scholar]
35. Emre M., Orgul S., Haufschild T., Shaw S.G., Flammer J. Increased plasma endothelin-1 levels in patients with progressive open angle glaucoma. Br. J. Ophthalmol. 2005;89:60–63. doi: 10.1136/bjo.2004.046755. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
36. McGrady N.R., Minton A.Z., Stankowska D.L., He S., Jefferies H.B., Krishnamoorthy R.R. Upregulation of the endothelin A (ETA) receptor and its association with neurodegeneration in a rodent model of glaucoma. BMC Neurosci. 2017;18:27. doi: 10.1186/s12868-017-0346-3. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
37. Lommatzsch C., Rothaus K., Schopmeyer L., Feldmann M., Bauer D., Grisanti S., Heinz C., Kasper M. Elevated endothelin-1 levels as risk factor for an impaired ocular blood flow measured by OCT-A in glaucoma. Sci. Rep. 2022;12:11801. doi: 10.1038/s41598-022-15401-5. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
38. Orgul S., Cioffi G.A., Bacon D.R., Van Buskirk E.M. An endothelin-1-induced model of chronic optic nerve ischemia in rhesus monkeys. J. Glaucoma. 1996;5:135–138. [PubMed] [Google Scholar]
39. Wang L., Fortune B., Cull G., Dong J., Cioffi G.A. Endothelin B receptor in human glaucoma and experimentally induced optic nerve damage. Arch. Ophthalmol. 2006;124:717–724. doi: 10.1001/archopht.124.5.717. [PubMed] [CrossRef] [Google Scholar]
40. Haefliger I.O., Dettmann E., Liu R., Meyer P., Prunte C., Messerli J., Flammer J. Potential role of nitric oxide and endothelin in the pathogenesis of glaucoma. Surv. Ophthalmol. 1999;43((Suppl. 1)):S51–S58. doi: 10.1016/S0039-6257(99)00026-0. [PubMed] [CrossRef] [Google Scholar]
41. Toda N., Nakanishi-Toda M. Nitric oxide: Ocular blood flow, glaucoma, and diabetic retinopathy. Prog. Retin. Eye Res. 2007;26:205–238. doi: 10.1016/j.preteyeres.2007.01.004. [PubMed] [CrossRef] [Google Scholar]
42. Masland R.H. The fundamental plan of the retina. Nat. Neurosci. 2001;4:877–886. doi: 10.1038/nn0901-877. [PubMed] [CrossRef] [Google Scholar]
43. Boccuni I., Fairless R. Retinal Glutamate Neurotransmission: From Physiology to Pathophysiological Mechanisms of Retinal Ganglion Cell Degeneration. Life. 2022;12:638. doi: 10.3390/life12050638. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
44. Lotery A.J. Glutamate excitotoxicity in glaucoma: Truth or fiction? Eye. 2005;19:369–370. doi: 10.1038/sj.eye.6701623. [PubMed] [CrossRef] [Google Scholar]
45. Inokuchi Y., Shimazawa M., Nakajima Y., Komuro I., Matsuda T., Baba A., Araie M., Kita S., Iwamoto T., Hara H. A Na+/Ca2+ exchanger isoform, NCX1, is involved in retinal cell death after N-methyl-D-aspartate injection and ischemia-reperfusion. J. Neurosci. Res. 2009;87:906–917. doi: 10.1002/jnr.21906. [PubMed] [CrossRef] [Google Scholar]
46. Brittain M.K., Brustovetsky T., Sheets P.L., Brittain J.M., Khanna R., Cummins T.R., Brustovetsky N. Delayed calcium dysregulation in neurons requires both the NMDA receptor and the reverse Na+/Ca2+ exchanger. Neurobiol. Dis. 2012;46:109–117. doi: 10.1016/j.nbd.2011.12.051. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
47. Brandt S.K., Weatherly M.E., Ware L., Linn D.M., Linn C.L. Calcium preconditioning triggers neuroprotection in retinal ganglion cells. Neuroscience. 2011;172:387–397. doi: 10.1016/j.neuroscience.2010.10.071. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
48. Hartwick A.T., Hamilton C.M., Baldridge W.H. Glutamatergic calcium dynamics and deregulation of rat retinal ganglion cells. J. Physiol. 2008;586:3425–3446. doi: 10.1113/jphysiol.2008.154609. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
49. Almasieh M., Wilson A.M., Morquette B., Cueva Vargas J.L., Di Polo A. The molecular basis of retinal ganglion cell death in glaucoma. Prog. Retin. Eye Res. 2012;31:152–181. doi: 10.1016/j.preteyeres.2011.11.002. [PubMed] [CrossRef] [Google Scholar]
50. Bringmann A., Pannicke T., Grosche J., Francke M., Wiedemann P., Skatchkov S.N., Osborne N.N., Reichenbach A. Muller cells in the healthy and diseased retina. Prog. Retin. Eye Res. 2006;25:397–424. doi: 10.1016/j.preteyeres.2006.05.003. [PubMed] [CrossRef] [Google Scholar]
51. Martin K.R., Levkovitch-Verbin H., Valenta D., Baumrind L., Pease M.E., Quigley H.A. Retinal glutamate transporter changes in experimental glaucoma and after optic nerve transection in the rat. Invest. Ophthalmol. Vis. Sci. 2002;43:2236–2243. [PubMed] [Google Scholar]
52. Schuettauf F., Thaler S., Bolz S., Fries J., Kalbacher H., Mankowska A., Zurakowski D., Zrenner E., Rejdak R. Alterations of amino acids and glutamate transport in the DBA/2J mouse retina; possible clues to degeneration. Graefes Arch. Clin. Exp. Ophthalmol. 2007;245:1157–1168. doi: 10.1007/s00417-006-0531-z. [PubMed] [CrossRef] [Google Scholar]
53. Harada T., Harada C., Nakamura K., Quah H.M., Okumura A., Namekata K., Saeki T., Aihara M., Yoshida H., Mitani A., et al. The potential role of glutamate transporters in the pathogenesis of normal tension glaucoma. J. Clin. Investig. 2007;117:1763–1770. doi: 10.1172/JCI30178. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
54. Johnson J.E., Barde Y.A., Schwab M., Thoenen H. Brain-derived neurotrophic factor supports the survival of cultured rat retinal ganglion cells. J. Neurosci. 1986;6:3031–3038. doi: 10.1523/JNEUROSCI.06-10-03031.1986. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
55. Dekeyster E., Geeraerts E., Buyens T., Van den Haute C., Baekelandt V., De Groef L., Salinas-Navarro M., Moons L. Tackling Glaucoma from within the Brain: An Unfortunate Interplay of BDNF and TrkB. PLoS ONE. 2015;10:e0142067. doi: 10.1371/journal.pone.0142067. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
56. Gonzalez-Hoyuela M., Barbas J.A., Rodriguez-Tebar A. The autoregulation of retinal ganglion cell number. Development. 2001;128:117–124. doi: 10.1242/dev.128.1.117. [PubMed] [CrossRef] [Google Scholar]
57. Kimura A., Namekata K., Guo X., Harada C., Harada T. Neuroprotection, Growth Factors and BDNF-TrkB Signalling in Retinal Degeneration. Int. J. Mol. Sci. 2016;17:1584. doi: 10.3390/ijms17091584. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
58. Lambuk L., Mohd Lazaldin M.A., Ahmad S., Iezhitsa I., Agarwal R., Uskokovic V., Mohamud R. Brain-Derived Neurotrophic Factor-Mediated Neuroprotection in Glaucoma: A Review of Current State of the Art. Front. Pharmacol. 2022;13:875662. doi: 10.3389/fphar.2022.875662. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
59. Pease M.E., McKinnon S.J., Quigley H.A., Kerrigan-Baumrind L.A., Zack D.J. Obstructed axonal transport of BDNF and its receptor TrkB in experimental glaucoma. Invest. Ophthalmol. Vis. Sci. 2000;41:764–774. [PubMed] [Google Scholar]
60. Quigley H.A., McKinnon S.J., Zack D.J., Pease M.E., Kerrigan-Baumrind L.A., Kerrigan D.F., Mitchell R.S. Retrograde axonal transport of BDNF in retinal ganglion cells is blocked by acute IOP elevation in rats. Invest. Ophthalmol. Vis. Sci. 2000;41:3460–3466. [PubMed] [Google Scholar]
61. Osborne A., Khatib T.Z., Songra L., Barber A.C., Hall K., Kong G.Y.X., Widdowson P.S., Martin K.R. Neuroprotection of retinal ganglion cells by a novel gene therapy construct that achieves sustained enhancement of brain-derived neurotrophic factor/tropomyosin-related kinase receptor-B signaling. Cell Death Dis. 2018;9:1007. doi: 10.1038/s41419-018-1041-8. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
62. Igarashi T., Nakamoto K., Kobayashi M., Suzuki H., Arima T., Tobita Y., Takao K., Igarashi T., Okuda T., Okada T., et al. Brain-derived Neurotrophic Factor in the Aqueous Humor of Glaucoma Patients. J. Nippon. Med. Sch. 2021;88:128–132. doi: 10.1272/jnms.JNMS.2021_88-305. [PubMed] [CrossRef] [Google Scholar]
63. Shpak A.A., Guekht A.B., Druzhkova T.A., Kozlova K.I., Gulyaeva N.V. Brain-Derived Neurotrophic Factor in Patients with Primary Open-Angle Glaucoma and Age-related Cataract. Curr. Eye Res. 2018;43:224–231. doi: 10.1080/02713683.2017.1396617. [PubMed] [CrossRef] [Google Scholar]
64. Tezel G. Multifactorial Pathogenic Processes of Retinal Ganglion Cell Degeneration in Glaucoma towards Multi-Target Strategies for Broader Treatment Effects. Cells. 2021;10:1372. doi: 10.3390/cells10061372. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
65. Osellame L.D., Blacker T.S., Duchen M.R. Cellular and molecular mechanisms of mitochondrial function. Best. Pract. Res. Clin. Endocrinol. Metab. 2012;26:711–723. doi: 10.1016/j.beem.2012.05.003. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
66. Halliwell B. Oxidative stress and neurodegeneration: Where are we now? J. Neurochem. 2006;97:1634–1658. doi: 10.1111/j.1471-4159.2006.03907.x. [PubMed] [CrossRef] [Google Scholar]
67. Izzotti A., Sacca S.C., Longobardi M., Cartiglia C. Sensitivity of ocular anterior chamber tissues to oxidative damage and its relevance to the pathogenesis of glaucoma. Invest. Ophthalmol. Vis. Sci. 2009;50:5251–5258. doi: 10.1167/iovs.09-3871. [PubMed] [CrossRef] [Google Scholar]
68. Clopton D.A., Saltman P. Low-level oxidative stress causes cell-cycle specific arrest in cultured cells. Biochem. Biophys. Res. Commun. 1995;210:189–196. doi: 10.1006/bbrc.1995.1645. [PubMed] [CrossRef] [Google Scholar]
69. Giancotti F.G. Integrin signaling: Specificity and control of cell survival and cell cycle progression. Curr. Opin. Cell Biol. 1997;9:691–700. doi: 10.1016/S0955-0674(97)80123-8. [PubMed] [CrossRef] [Google Scholar]
70. Knepper P.A., Goossens W., Hvizd M., Palmberg P.F. Glycosaminoglycans of the human trabecular meshwork in primary open-angle glaucoma. Invest. Ophthalmol. Vis. Sci. 1996;37:1360–1367. [PubMed] [Google Scholar]
71. Zhou L., Li Y., Yue B.Y. Oxidative stress affects cytoskeletal structure and cell-matrix interactions in cells from an ocular tissue: The trabecular meshwork. J. Cell Physiol. 1999;180:182–189. doi: 10.1002/(SICI)1097-4652(199908)180:2<182::AID-JCP6>3.0.CO;2-X. [PubMed] [CrossRef] [Google Scholar]
72. Alvarado J.A., Alvarado R.G., Yeh R.F., Franse-Carman L., Marcellino G.R., Brownstein M.J. A new insight into the cellular regulation of aqueous outflow: How trabecular meshwork endothelial cells drive a mechanism that regulates the permeability of Schlemm’s canal endothelial cells. Br. J. Ophthalmol. 2005;89:1500–1505. doi: 10.1136/bjo.2005.081307. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
73. Kong G.Y.X., Van Bergen N.J., Trounce I.A., Crowston J.G. Mitochondrial Dysfunction and Glaucoma. J. Glaucoma. 2009;18:93–100. doi: 10.1097/IJG.0b013e318181284f. [PubMed] [CrossRef] [Google Scholar]
74. He Y., Leung K.W., Zhang Y.H., Duan S., Zhong X.F., Jiang R.Z., Peng Z., Tombran-Tink J., Ge J. Mitochondrial complex I defect induces ROS release and degeneration in trabecular meshwork cells of POAG patients: Protection by antioxidants. Invest. Ophthalmol. Vis. Sci. 2008;49:1447–1458. doi: 10.1167/iovs.07-1361. [PubMed] [CrossRef] [Google Scholar]
75. Mittag T.W., Danias J., Pohorenec G., Yuan H.M., Burakgazi E., Chalmers-Redman R., Podos S.M., Tatton W.G. Retinal damage after 3 to 4 months of elevated intraocular pressure in a rat glaucoma model. Invest. Ophthalmol. Vis. Sci. 2000;41:3451–3459. [PubMed] [Google Scholar]
76. Shi W., Tan C., Liu C., Chen D. Mitochondrial fission mediated by Drp1-Fis1 pathway and neurodegenerative diseases. Rev. Neurosci. 2023;34:275–294. doi: 10.1515/revneuro-2022-0056. [PubMed] [CrossRef] [Google Scholar]
77. Zeng Z., You M., Fan C., Rong R., Li H., Xia X. Pathologically high intraocular pressure induces mitochondrial dysfunction through Drp1 and leads to retinal ganglion cell PANoptosis in glaucoma. Redox Biol. 2023;62:102687. doi: 10.1016/j.redox.2023.102687. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
78. Tezel G. Molecular regulation of neuroinflammation in glaucoma: Current knowledge and the ongoing search for new treatment targets. Prog. Retin. Eye Res. 2022;87:100998. doi: 10.1016/j.preteyeres.2021.100998. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
79. Kaur C., Ling E.A. Blood brain barrier in hypoxic-ischemic conditions. Curr. Neurovasc Res. 2008;5:71–81. doi: 10.2174/156720208783565645. [PubMed] [CrossRef] [Google Scholar]
80. Chua J., Vania M., Cheung C.M., Ang M., Chee S.P., Yang H., Li J., Wong T.T. Expression profile of inflammatory cytokines in aqueous from glaucomatous eyes. Mol. Vis. 2012;18:431–438. [PMC free article] [PubMed] [Google Scholar]
81. Wei X., Cho K.S., Thee E.F., Jager M.J., Chen D.F. Neuroinflammation and microglia in glaucoma: Time for a paradigm shift. J. Neurosci. Res. 2019;97:70–76. doi: 10.1002/jnr.24256. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
82. Howell G.R., Macalinao D.G., Sousa G.L., Walden M., Soto I., Kneeland S.C., Barbay J.M., King B.L., Marchant J.K., Hibbs M., et al. Molecular clustering identifies complement and endothelin induction as early events in a mouse model of glaucoma. J. Clin. Investig. 2011;121:1429–1444. doi: 10.1172/JCI44646. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
83. Chen H., Cho K.S., Vu T.H.K., Shen C.H., Kaur M., Chen G., Mathew R., McHam M.L., Fazelat A., Lashkari K., et al. Commensal microflora-induced T cell responses mediate progressive neurodegeneration in glaucoma. Nat. Commun. 2018;9:3209. doi: 10.1038/s41467-018-05681-9. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
84. Von Thun Und Hohenstein-Blaul N., Bell K., Pfeiffer N., Grus F.H. Autoimmune aspects in glaucoma. Eur. J. Pharmacol. 2016;787:105–118. doi: 10.1016/j.ejphar.2016.04.031. [PubMed] [CrossRef] [Google Scholar]
85. Yuan L., Neufeld A.H. Activated microglia in the human glaucomatous optic nerve head. J. Neurosci. Res. 2001;64:523–532. doi: 10.1002/jnr.1104. [PubMed] [CrossRef] [Google Scholar]
86. de Hoz R., Gallego B.I., Ramirez A.I., Rojas B., Salazar J.J., Valiente-Soriano F.J., Aviles-Trigueros M., Villegas-Perez M.P., Vidal-Sanz M., Trivino A., et al. Rod-like microglia are restricted to eyes with laser-induced ocular hypertension but absent from the microglial changes in the contralateral untreated eye. PLoS ONE. 2013;8:e83733. doi: 10.1371/journal.pone.0083733. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
87. Huang D., Swanson E.A., Lin C.P., Schuman J.S., Stinson W.G., Chang W., Hee M.R., Flotte T., Gregory K., Puliafito C.A., et al. Optical coherence tomography. Science. 1991;254:1178–1181. doi: 10.1126/science.1957169. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
88. Drexler W., Fujimoto J.G. State-of-the-art retinal optical coherence tomography. Prog. Retin. Eye Res. 2008;27:45–88. doi: 10.1016/j.preteyeres.2007.07.005. [PubMed] [CrossRef] [Google Scholar]
89. Geevarghese A., Wollstein G., Ishikawa H., Schuman J.S. Optical Coherence Tomography and Glaucoma. Annu. Rev. Vis. Sci. 2021;7:693–726. doi: 10.1146/annurev-vision-100419-111350. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
90. Sommer A., Katz J., Quigley H.A., Miller N.R., Robin A.L., Richter R.C., Witt K.A. Clinically detectable nerve fiber atrophy precedes the onset of glaucomatous field loss. Arch. Ophthalmol. 1991;109:77–83. doi: 10.1001/archopht.1991.01080010079037. [PubMed] [CrossRef] [Google Scholar]
91. Werkmeister R.M., Cherecheanu A.P., Garhofer G., Schmidl D., Schmetterer L. Imaging of retinal ganglion cells in glaucoma: Pitfalls and challenges. Cell Tissue Res. 2013;353:261–268. doi: 10.1007/s00441-013-1600-3. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
92. Dong Z.M., Wollstein G., Wang B., Schuman J.S. Adaptive optics optical coherence tomography in glaucoma. Prog. Retin. Eye Res. 2017;57:76–88. doi: 10.1016/j.preteyeres.2016.11.001. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
93. Kocaoglu O.P., Cense B., Jonnal R.S., Wang Q., Lee S., Gao W., Miller D.T. Imaging retinal nerve fiber bundles using optical coherence tomography with adaptive optics. Vision. Res. 2011;51:1835–1844. doi: 10.1016/j.visres.2011.06.013. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
94. Sarossy M., Crowston J., Kumar D., Weymouth A., Wu Z. Prediction of glaucoma severity using parameters from the electroretinogram. Sci. Rep. 2021;11:23886. doi: 10.1038/s41598-021-03421-6. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
95. Banitt M.R., Ventura L.M., Feuer W.J., Savatovsky E., Luna G., Shif O., Bosse B., Porciatti V. Progressive loss of retinal ganglion cell function precedes structural loss by several years in glaucoma suspects. Invest. Ophthalmol. Vis. Sci. 2013;54:2346–2352. doi: 10.1167/iovs.12-11026. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
96. Bode S.F., Jehle T., Bach M. Pattern electroretinogram in glaucoma suspects: New findings from a longitudinal study. Invest. Ophthalmol. Vis. Sci. 2011;52:4300–4306. doi: 10.1167/iovs.10-6381. [PubMed] [CrossRef] [Google Scholar]
97. Jung K.I., Jeon S., Shin D.Y., Lee J., Park C.K. Pattern Electroretinograms in Preperimetric and Perimetric Glaucoma. Am. J. Ophthalmol. 2020;215:118–126. doi: 10.1016/j.ajo.2020.02.008. [PubMed] [CrossRef] [Google Scholar]
98. Lee T., Seo D.R., Kim J.Y., Choi W., Lee S.Y., Lee J.M., Seong G.J., Kim C.Y., Bae H.W. Relationship between N95 Amplitude of Pattern Electroretinogram and Optical Coherence Tomography Angiography in Open-Angle Glaucoma. J. Clin. Med. 2020;9:3854. doi: 10.3390/jcm9123854. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
99. Viswanathan S., Frishman L.J., Robson J.G., Harwerth R.S., Smith E.L., 3rd The photopic negative response of the macaque electroretinogram: Reduction by experimental glaucoma. Invest. Ophthalmol. Vis. Sci. 1999;40:1124–1136. [PubMed] [Google Scholar]
100. Machida S., Gotoh Y., Toba Y., Ohtaki A., Kaneko M., Kurosaka D. Correlation between photopic negative response and retinal nerve fiber layer thickness and optic disc topography in glaucomatous eyes. Invest. Ophthalmol. Vis. Sci. 2008;49:2201–2207. doi: 10.1167/iovs.07-0887. [PubMed] [CrossRef] [Google Scholar]
101. Wilsey L.J., Fortune B. Electroretinography in glaucoma diagnosis. Curr. Opin. Ophthalmol. 2016;27:118–124. doi: 10.1097/ICU.0000000000000241. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
102. Chan H.L., Brown B. Multifocal ERG changes in glaucoma. Ophthalmic Physiol. Opt. 1999;19:306–316. doi: 10.1111/j.1475-1313.1999.00439.x. [PubMed] [CrossRef] [Google Scholar]
103. Hasegawa S., Takagi M., Usui T., Takada R., Abe H. Waveform changes of the first-order multifocal electroretinogram in patients with glaucoma. Invest. Ophthalmol. Vis. Sci. 2000;41:1597–1603. [PubMed] [Google Scholar]
104. Fortune B., Bearse M.A., Jr., Cioffi G.A., Johnson C.A. Selective loss of an oscillatory component from temporal retinal multifocal ERG responses in glaucoma. Invest. Ophthalmol. Vis. Sci. 2002;43:2638–2647. [PubMed] [Google Scholar]
105. Palmowski A.M., Allgayer R., Heinemann-Vemaleken B. The multifocal ERG in open angle glaucoma--a comparison of high and low contrast recordings in high- and low-tension open angle glaucoma. Doc. Ophthalmol. 2000;101:35–49. doi: 10.1023/A:1002710707763. [PubMed] [CrossRef] [Google Scholar]
106. Gardiner S.K., Swanson W.H., Demirel S., McKendrick A.M., Turpin A., Johnson C.A. A two-stage neural spiking model of visual contrast detection in perimetry. Vision. Res. 2008;48:1859–1869. doi: 10.1016/j.visres.2008.06.006. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
107. Dacey D.M., Lee B.B. The ‘blue-on’ opponent pathway in primate retina originates from a distinct bistratified ganglion cell type. Nature. 1994;367:731–735. doi: 10.1038/367731a0. [PubMed] [CrossRef] [Google Scholar]
108. Sample P.A., Bosworth C.F., Blumenthal E.Z., Girkin C., Weinreb R.N. Visual function-specific perimetry for indirect comparison of different ganglion cell populations in glaucoma. Invest. Ophthalmol. Vis. Sci. 2000;41:1783–1790. [PubMed] [Google Scholar]
109. Johnson C.A., Adams A.J., Casson E.J., Brandt J.D. Progression of early glaucomatous visual field loss as detected by blue-on-yellow and standard white-on-white automated perimetry. Arch. Ophthalmol. 1993;111:651–656. doi: 10.1001/archopht.1993.01090050085035. [PubMed] [CrossRef] [Google Scholar]
110. Demirel S., Johnson C.A. Incidence and prevalence of short wavelength automated perimetry deficits in ocular hypertensive patients. Am. J. Ophthalmol. 2001;131:709–715. doi: 10.1016/S0002-9394(00)00946-6. [PubMed] [CrossRef] [Google Scholar]
111. Medeiros F.A., Sample P.A., Weinreb R.N. Corneal thickness measurements and visual function abnormalities in ocular hypertensive patients. Am. J. Ophthalmol. 2003;135:131–137. doi: 10.1016/S0002-9394(02)01886-X. [PubMed] [CrossRef] [Google Scholar]
112. Shah N.N., Bowd C., Medeiros F.A., Weinreb R.N., Sample P.A., Hoffmann E.M., Zangwill L.M. Combining structural and functional testing for detection of glaucoma. Ophthalmology. 2006;113:1593–1602. doi: 10.1016/j.ophtha.2006.06.004. [PubMed] [CrossRef] [Google Scholar]
113. Medeiros F.A., Sample P.A., Weinreb R.N. Frequency doubling technology perimetry abnormalities as predictors of glaucomatous visual field loss. Am. J. Ophthalmol. 2004;137:863–871. doi: 10.1016/j.ajo.2003.12.009. [PubMed] [CrossRef] [Google Scholar]
114. Johnson C.A., Samuels S.J. Screening for glaucomatous visual field loss with frequency-doubling perimetry. Invest. Ophthalmol. Vis. Sci. 1997;38:413–425. [PubMed] [Google Scholar]
115. Chauhan B.C., Johnson C.A. Test-retest variability of frequency-doubling perimetry and conventional perimetry in glaucoma patients and normal subjects. Invest. Ophthalmol. Vis. Sci. 1999;40:648–656. [PubMed] [Google Scholar]
116. Cordeiro M.F., Hill D., Patel R., Corazza P., Maddison J., Younis S. Detecting retinal cell stress and apoptosis with DARC: Progression from lab to clinic. Prog. Retin. Eye Res. 2022;86:100976. doi: 10.1016/j.preteyeres.2021.100976. [PubMed] [CrossRef] [Google Scholar]
117. Cordeiro M.F., Guo L., Luong V., Harding G., Wang W., Jones H.E., Moss S.E., Sillito A.M., Fitzke F.W. Real-time imaging of single nerve cell apoptosis in retinal neurodegeneration. Proc. Natl. Acad. Sci. USA. 2004;101:13352–13356. doi: 10.1073/pnas.0405479101. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
118. Guo L., Moss S.E., Alexander R.A., Ali R.R., Fitzke F.W., Cordeiro M.F. Retinal ganglion cell apoptosis in glaucoma is related to intraocular pressure and IOP-induced effects on extracellular matrix. Invest. Ophthalmol. Vis. Sci. 2005;46:175–182. doi: 10.1167/iovs.04-0832. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
119. Yoles E., Wheeler L.A., Schwartz M. Alpha2-adrenoreceptor agonists are neuroprotective in a rat model of optic nerve degeneration. Invest. Ophthalmol. Vis. Sci. 1999;40:65–73. [PubMed] [Google Scholar]
120. Kalapesi F.B., Coroneo M.T., Hill M.A. Human ganglion cells express the alpha-2 adrenergic receptor: Relevance to neuroprotection. Br. J. Ophthalmol. 2005;89:758–763. doi: 10.1136/bjo.2004.053025. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
121. Prokosch V., Panagis L., Volk G.F., Dermon C., Thanos S. Alpha2-adrenergic receptors and their core involvement in the process of axonal growth in retinal explants. Invest. Ophthalmol. Vis. Sci. 2010;51:6688–6699. doi: 10.1167/iovs.09-4835. [PubMed] [CrossRef] [Google Scholar]
122. Lambert W.S., Ruiz L., Crish S.D., Wheeler L.A., Calkins D.J. Brimonidine prevents axonal and somatic degeneration of retinal ganglion cell neurons. Mol. Neurodegener. 2011;6:4. doi: 10.1186/1750-1326-6-4. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
123. Krupin T., Liebmann J.M., Greenfield D.S., Ritch R., Gardiner S., Low-Pressure Glaucoma Study G. A randomized trial of brimonidine versus timolol in preserving visual function: Results from the Low-Pressure Glaucoma Treatment Study. Am. J. Ophthalmol. 2011;151:671–681. doi: 10.1016/j.ajo.2010.09.026. [PubMed] [CrossRef] [Google Scholar]
124. Tsai J.C., Chang H.W. Comparison of the effects of brimonidine 0.2% and timolol 0.5% on retinal nerve fiber layer thickness in ocular hypertensive patients: A prospective, unmasked study. J. Ocul. Pharmacol. Ther. 2005;21:475–482. doi: 10.1089/jop.2005.21.475. [PubMed] [CrossRef] [Google Scholar]
125. Guo L., Salt T.E., Maass A., Luong V., Moss S.E., Fitzke F.W., Cordeiro M.F. Assessment of neuroprotective effects of glutamate modulation on glaucoma-related retinal ganglion cell apoptosis in vivo. Invest. Ophthalmol. Vis. Sci. 2006;47:626–633. doi: 10.1167/iovs.05-0754. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
126. Olney J.W., Labruyere J., Wang G., Wozniak D.F., Price M.T., Sesma M.A. NMDA antagonist neurotoxicity: Mechanism and prevention. Science. 1991;254:1515–1518. doi: 10.1126/science.1835799. [PubMed] [CrossRef] [Google Scholar]
127. Hare W.A., WoldeMussie E., Lai R.K., Ton H., Ruiz G., Chun T., Wheeler L. Efficacy and safety of memantine treatment for reduction of changes associated with experimental glaucoma in monkey, I: Functional measures. Invest. Ophthalmol. Vis. Sci. 2004;45:2625–2639. doi: 10.1167/iovs.03-0566. [PubMed] [CrossRef] [Google Scholar]
128. Vorwerk C.K., Lipton S.A., Zurakowski D., Hyman B.T., Sabel B.A., Dreyer E.B. Chronic low-dose glutamate is toxic to retinal ganglion cells. Toxicity blocked by memantine. Invest. Ophthalmol. Vis. Sci. 1996;37:1618–1624. [PubMed] [Google Scholar]
129. Danesh-Meyer H.V., Levin L.A. Neuroprotection: Extrapolating from neurologic diseases to the eye. Am. J. Ophthalmol. 2009;148:186–191.e182. doi: 10.1016/j.ajo.2009.03.029. [PubMed] [CrossRef] [Google Scholar]
130. Osborne N.N., Wood J.P., Cupido A., Melena J., Chidlow G. Topical flunarizine reduces IOP and protects the retina against ischemia-excitotoxicity. Invest. Ophthalmol. Vis. Sci. 2002;43:1456–1464. [PubMed] [Google Scholar]
131. Koseki N., Araie M., Tomidokoro A., Nagahara M., Hasegawa T., Tamaki Y., Yamamoto S. A placebo-controlled 3-year study of a calcium blocker on visual field and ocular circulation in glaucoma with low-normal pressure. Ophthalmology. 2008;115:2049–2057. doi: 10.1016/j.ophtha.2008.05.015. [PubMed] [CrossRef] [Google Scholar]
132. Sawada A., Kitazawa Y., Yamamoto T., Okabe I., Ichien K. Prevention of visual field defect progression with brovincamine in eyes with normal-tension glaucoma. Ophthalmology. 1996;103:283–288. doi: 10.1016/S0161-6420(96)30703-3. [PubMed] [CrossRef] [Google Scholar]
133. Nucci C., Tartaglione R., Cerulli A., Mancino R., Spano A., Cavaliere F., Rombola L., Bagetta G., Corasaniti M.T., Morrone L.A. Retinal damage caused by high intraocular pressure-induced transient ischemia is prevented by coenzyme Q10 in rat. Int. Rev. Neurobiol. 2007;82:397–406. doi: 10.1016/S0074-7742(07)82022-8. [PubMed] [CrossRef] [Google Scholar]
134. Russo R., Cavaliere F., Rombola L., Gliozzi M., Cerulli A., Nucci C., Fazzi E., Bagetta G., Corasaniti M.T., Morrone L.A. Rational basis for the development of coenzyme Q10 as a neurotherapeutic agent for retinal protection. Prog. Brain Res. 2008;173:575–582. doi: 10.1016/S0079-6123(08)01139-4. [PubMed] [CrossRef] [Google Scholar]
135. Nakajima Y., Inokuchi Y., Nishi M., Shimazawa M., Otsubo K., Hara H. Coenzyme Q10 protects retinal cells against oxidative stress in vitro and in vivo. Brain Res. 2008;1226:226–233. doi: 10.1016/j.brainres.2008.06.026. [PubMed] [CrossRef] [Google Scholar]
136. Lee D., Shim M.S., Kim K.Y., Noh Y.H., Kim H., Kim S.Y., Weinreb R.N., Ju W.K. Coenzyme Q10 inhibits glutamate excitotoxicity and oxidative stress-mediated mitochondrial alteration in a mouse model of glaucoma. Invest. Ophthalmol. Vis. Sci. 2014;55:993–1005. doi: 10.1167/iovs.13-12564. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
137. Parisi V., Centofanti M., Gandolfi S., Marangoni D., Rossetti L., Tanga L., Tardini M., Traina S., Ungaro N., Vetrugno M., et al. Effects of coenzyme Q10 in conjunction with vitamin E on retinal-evoked and cortical-evoked responses in patients with open-angle glaucoma. J. Glaucoma. 2014;23:391–404. doi: 10.1097/IJG.0b013e318279b836. [PubMed] [CrossRef] [Google Scholar]
138. Sawai H., Clarke D.B., Kittlerova P., Bray G.M., Aguayo A.J. Brain-derived neurotrophic factor and neurotrophin-4/5 stimulate growth of axonal branches from regenerating retinal ganglion cells. J. Neurosci. 1996;16:3887–3894. doi: 10.1523/JNEUROSCI.16-12-03887.1996. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
139. Chen H., Weber A.J. BDNF enhances retinal ganglion cell survival in cats with optic nerve damage. Invest. Ophthalmol. Vis. Sci. 2001;42:966–974. [PubMed] [Google Scholar]
140. Domenici L., Origlia N., Falsini B., Cerri E., Barloscio D., Fabiani C., Sansò M., Giovannini L. Rescue of Retinal Function by BDNF in a Mouse Model of Glaucoma. PLoS ONE. 2014;9:e115579. doi: 10.1371/journal.pone.0115579. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
141. Ko M.L., Hu D.N., Ritch R., Sharma S.C., Chen C.F. Patterns of retinal ganglion cell survival after brain-derived neurotrophic factor administration in hypertensive eyes of rats. Neurosci. Lett. 2001;305:139–142. doi: 10.1016/S0304-3940(01)01830-4. [PubMed] [CrossRef] [Google Scholar]
142. Chen H., Weber A.J. Brain-derived neurotrophic factor reduces TrkB protein and mRNA in the normal retina and following optic nerve crush in adult rats. Brain Res. 2004;1011:99–106. doi: 10.1016/j.brainres.2004.03.024. [PubMed] [CrossRef] [Google Scholar]
143. Kirsch M., Lee M.Y., Meyer V., Wiese A., Hofmann H.D. Evidence for multiple, local functions of ciliary neurotrophic factor (CNTF) in retinal development: Expression of CNTF and its receptors and in vitro effects on target cells. J. Neurochem. 1997;68:979–990. doi: 10.1046/j.1471-4159.1997.68030979.x. [PubMed] [CrossRef] [Google Scholar]
144. Wen R., Song Y., Liu Y., Li Y., Zhao L., Laties A.M. CNTF negatively regulates the phototransduction machinery in rod photoreceptors: Implication for light-induced photostasis plasticity. Adv. Exp. Med. Biol. 2008;613:407–413. doi: 10.1007/978-0-387-74904-4_48. [PubMed] [CrossRef] [Google Scholar]
145. Mey J., Thanos S. Intravitreal injections of neurotrophic factors support the survival of axotomized retinal ganglion cells in adult rats in vivo. Brain Res. 1993;602:304–317. doi: 10.1016/0006-8993(93)90695-J. [PubMed] [CrossRef] [Google Scholar]
146. Takahata K., Katsuki H., Kume T., Nakata D., Ito K., Muraoka S., Yoneda F., Kashii S., Honda Y., Akaike A. Retinal neuronal death induced by intraocular administration of a nitric oxide donor and its rescue by neurotrophic factors in rats. Invest. Ophthalmol. Vis. Sci. 2003;44:1760–1766. doi: 10.1167/iovs.02-0471. [PubMed] [CrossRef] [Google Scholar]
147. Colafrancesco V., Parisi V., Sposato V., Rossi S., Russo M.A., Coassin M., Lambiase A., Aloe L. Ocular application of nerve growth factor protects degenerating retinal ganglion cells in a rat model of glaucoma. J. Glaucoma. 2011;20:100–108. doi: 10.1097/IJG.0b013e3181d787e5. [PubMed] [CrossRef] [Google Scholar]
148. Lambiase A., Aloe L., Centofanti M., Parisi V., Mantelli F., Colafrancesco V., Manni G.L., Bucci M.G., Bonini S., Levi-Montalcini R. Experimental and clinical evidence of neuroprotection by nerve growth factor eye drops: Implications for glaucoma. Proc. Natl. Acad. Sci. USA. 2009;106:13469–13474. doi: 10.1073/pnas.0906678106. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
149. Guo L., Davis B.M., Ravindran N., Galvao J., Kapoor N., Haamedi N., Shamsher E., Luong V., Fico E., Cordeiro M.F. Topical recombinant human Nerve growth factor (rh-NGF) is neuroprotective to retinal ganglion cells by targeting secondary degeneration. Sci. Rep. 2020;10:3375. doi: 10.1038/s41598-020-60427-2. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
150. Jiang C., Moore M.J., Zhang X., Klassen H., Langer R., Young M. Intravitreal injections of GDNF-loaded biodegradable microspheres are neuroprotective in a rat model of glaucoma. Mol. Vis. 2007;13:1783–1792. [PubMed] [Google Scholar]
151. Checa-Casalengua P., Jiang C., Bravo-Osuna I., Tucker B.A., Molina-Martínez I.T., Young M.J., Herrero-Vanrell R. Retinal ganglion cells survival in a glaucoma model by GDNF/Vit E PLGA microspheres prepared according to a novel microencapsulation procedure. J. Control. Release. 2011;156:92–100. doi: 10.1016/j.jconrel.2011.06.023. [PubMed] [CrossRef] [Google Scholar]
152. Ward M., Khoobehi A., Lavik E., Langer R., Young M. Neuroprotection of retinal ganglion cells in DBA/2J mice with GDNF-loaded biodegradable microspheres. J. Pharm. Sci. 2007;96:558–568. doi: 10.1002/jps.20629. [PubMed] [CrossRef] [Google Scholar]
153. Wang Q., Zhuang P., Huang H., Li L., Liu L., Webber H.C., Dalal R., Siew L., Fligor C.M., Chang K.C., et al. Mouse γ-Synuclein Promoter-Mediated Gene Expression and Editing in Mammalian Retinal Ganglion Cells. J. Neurosci. 2020;40:3896–3914. doi: 10.1523/JNEUROSCI.0102-20.2020. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
154. Bosco A., Anderson S.R., Breen K.T., Romero C.O., Steele M.R., Chiodo V.A., Boye S.L., Hauswirth W.W., Tomlinson S., Vetter M.L. Complement C3-Targeted Gene Therapy Restricts Onset and Progression of Neurodegeneration in Chronic Mouse Glaucoma. Mol. Ther. 2018;26:2379–2396. doi: 10.1016/j.ymthe.2018.08.017. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
155. Guo X., Zhou J., Starr C., Mohns E.J., Li Y., Chen E.P., Yoon Y., Kellner C.P., Tanaka K., Wang H., et al. Preservation of vision after CaMKII-mediated protection of retinal ganglion cells. Cell. 2021;184:4299–4314.e4212. doi: 10.1016/j.cell.2021.06.031. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
156. Fang F., Zhuang P., Feng X., Liu P., Liu D., Huang H., Li L., Chen W., Liu L., Sun Y., et al. NMNAT2 is downregulated in glaucomatous RGCs, and RGC-specific gene therapy rescues neurodegeneration and visual function. Mol. Ther. 2022;30:1421–1431. doi: 10.1016/j.ymthe.2022.01.035. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
157. Visuvanathan S., Baker A.N., Lagali P.S., Coupland S.G., Miller G., Hauswirth W.W., Tsilfidis C. XIAP gene therapy effects on retinal ganglion cell structure and function in a mouse model of glaucoma. Gene Ther. 2022;29:147–156. doi: 10.1038/s41434-021-00281-7. [PubMed] [CrossRef] [Google Scholar]
158. Donahue R.J., Fehrman R.L., Gustafson J.R., Nickells R.W. BCLXL gene therapy moderates neuropathology in the DBA/2J mouse model of inherited glaucoma. Cell Death Dis. 2021;12:781. doi: 10.1038/s41419-021-04068-x. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
159. Nishida A., Takahashi M., Tanihara H., Nakano I., Takahashi J.B., Mizoguchi A., Ide C., Honda Y. Incorporation and differentiation of hippocampus-derived neural stem cells transplanted in injured adult rat retina. Invest. Ophthalmol. Vis. Sci. 2000;41:4268–4274. [PubMed] [Google Scholar]
160. Singhal S., Bhatia B., Jayaram H., Becker S., Jones M.F., Cottrill P.B., Khaw P.T., Salt T.E., Limb G.A. Human Muller glia with stem cell characteristics differentiate into retinal ganglion cell (RGC) precursors in vitro and partially restore RGC function in vivo following transplantation. Stem Cells Transl. Med. 2012;1:188–199. doi: 10.5966/sctm.2011-0005. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
161. Wheeler L.A., Gil D.W., WoldeMussie E. Role of alpha-2 adrenergic receptors in neuroprotection and glaucoma. Surv. Ophthalmol. 2001;45((Suppl. 3)):S290–S294; discussion S295–S296. doi: 10.1016/S0039-6257(01)00206-5. [PubMed] [CrossRef] [Google Scholar]
162. Bylund D.B., Chacko D.M. Characterization of alpha2 adrenergic receptor subtypes in human ocular tissue homogenates. Invest. Ophthalmol. Vis. Sci. 1999;40:2299–2306. [PubMed] [Google Scholar]
163. Takayama J., Tomidokoro A., Ishii K., Tamaki Y., Fukaya Y., Hosokawa T., Araie M. Time course of the change in optic nerve head circulation after an acute increase in intraocular pressure. Invest. Ophthalmol. Vis. Sci. 2003;44:3977–3985. doi: 10.1167/iovs.03-0024. [PubMed] [CrossRef] [Google Scholar]
164. Harada C., Harada T., Quah H.-M., Maekawa F., Yoshida K., Ohno S., Wada K., Parada L., Tanaka K. Potential role of glial cell line-derived neurotrophic factor receptors in Müller glial cells during light-induced retinal degeneration. Neuroscience. 2003;122:229–235. doi: 10.1016/S0306-4522(03)00599-2. [PubMed] [CrossRef] [Google Scholar]
165. Ratican S.E., Osborne A., Martin K.R. Progress in Gene Therapy to Prevent Retinal Ganglion Cell Loss in Glaucoma and Leber’s Hereditary Optic Neuropathy. Neural Plast. 2018;2018:7108948. doi: 10.1155/2018/7108948. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
166. Conlon R., Saheb H., Ahmed I.I.K. Glaucoma treatment trends: A review. Can. J. Ophthalmol. 2017;52:114–124. doi: 10.1016/j.jcjo.2016.07.013. [PubMed] [CrossRef] [Google Scholar]
167. Kang J.M., Lin S. Ginkgo biloba and its potential role in glaucoma. Curr. Opin. Ophthalmol. 2018;29:116–120. doi: 10.1097/ICU.0000000000000459. [PubMed] [CrossRef] [Google Scholar]
168. Hirooka K., Tokuda M., Miyamoto O., Itano T., Baba T., Shiraga F. The Ginkgo biloba extract (EGb 761) provides a neuroprotective effect on retinal ganglion cells in a rat model of chronic glaucoma. Curr. Eye Res. 2004;28:153–157. doi: 10.1076/ceyr.28.3.153.26246. [PubMed] [CrossRef] [Google Scholar]
169. Ullah A., Munir S., Badshah S.L., Khan N., Ghani L., Poulson B.G., Emwas A.H., Jaremko M. Important Flavonoids and Their Role as a Therapeutic Agent. Molecules. 2020;25:5243. doi: 10.3390/molecules25225243. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
170. Luis J., Eastlake K., Lamb W.D.B., Limb G.A., Jayaram H., Khaw P.T. Cell-Based Therapies for Glaucoma. Transl. Vision. Sci. Technol. 2023;12:23. doi: 10.1167/tvst.12.7.23. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
171. Sahle F.F., Kim S., Niloy K.K., Tahia F., Fili C.V., Cooper E., Hamilton D.J., Lowe T.L. Nanotechnology in regenerative ophthalmology. Adv. Drug Deliv. Rev. 2019;148:290–307. doi: 10.1016/j.addr.2019.10.006. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
172. Yousefi S. Clinical Applications of Artificial Intelligence in Glaucoma. J. Ophthalmic Vis. Res. 2023;18:97–112. doi: 10.18502/jovr.v18i1.12730. [PMC free article] [PubMed] [CrossRef] [Google Scholar]