https://pmc.ncbi.nlm.nih.gov/articles/PMC4275385/

Review
Microbial short-chain fatty acids: a strategy to tune adoptive T cell therapy
- Priya Rangan and
- http://orcid.org/0000-0003-0833-6927Anna Mondino
- Correspondence to Dr Anna Mondino; mondino.anna@hsr.it
Abstract
The gut microbiota and its metabolites have been shown to play a pivotal role in the regulation of metabolic, endocrine and immune functions. Though the exact mechanism of action remains to be fully elucidated, available knowledge supports the ability of microbiota-fermented short-chain fatty acids (SCFAs), such as acetate, propionate, and butyrate, to influence epigenetic and metabolic cascades controlling gene expression, chemotaxis, differentiation, proliferation, and apoptosis in several non-immune and immune cell subsets. While used as preferred metabolic substrates and sources of energy by colonic gut epithelial cells, most recent evidence indicates that these metabolites regulate immune functions, and in particular fine-tune T cell effector, regulatory and memory phenotypes, with direct in vivo consequences on the efficacy of chemotherapy, radiotherapy and immunotherapy. Most recent data also support the use of these metabolites over the course of T cell manufacturing, paving the way for refined adoptive T cell therapy engineering. Here, we review the most recent advances in the field, highlighting in vitro and in vivo evidence for the ability of SCFAs to shape T cell phenotypes and functions.
http://creativecommons.org/licenses/by-nc/4.0/
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See http://creativecommons.org/licenses/by-nc/4.0/.
초록
장내 미생물군집과 그 대사산물은
대사, 내분비 및 면역 기능 조절에 핵심적인 역할을 한다는 것이 밝혀졌습니다.
정확한 작용 메커니즘은 아직 완전히 규명되지 않았지만,
현재까지의 연구 결과는 장내 미생물이 발효한 단쇄 지방산(SCFAs)인
아세테이트, 프로피오네이트, 부티레이트 등이
유전자 발현, 화학유도, 분화, 증식, 사멸을 조절하는
에피제네틱 및 대사 경로를 영향을 미칠 수 있음을 보여줍니다.
대장 장 상피 세포의 선호되는 대사 기질 및 에너지 공급원으로 사용되는 이 대사산물은
최근 연구에서 면역 기능을 조절하며,
특히 T 세포의 효과기, 조절 및 기억 형질을 미세 조정하여
화학요법, 방사선 요법 및 면역요법의 효능에 직접적인 영향을 미친다는 것이 밝혀졌습니다.
최근 데이터는
T 세포 제조 과정 전반에 걸쳐 이러한 대사 산물의 사용을 지원하며,
정교한 채택 T 세포 치료 공학의 길을 열었습니다.
본 논문에서는
SCFAs가 T 세포 표현형 및 기능을 형성하는 능력에 대한 체외 및 체내 증거를 강조하며,
해당 분야의 최신 진전을 검토합니다.
http://creativecommons.org/licenses/by-nc/4.0/
이 논문은 크리에이티브 커먼즈 Attribution Non Commercial (CC BY-NC 4.0) 라이선스에 따라 배포되며, 이는 원작을 적절히 인용하고 변경 사항을 표시하며 상업적 목적이 아닌 경우에 한해 배포, 재혼합, 적응, 파생 작품 제작을 허용합니다. 자세한 내용은 http://creativecommons.org/licenses/by-nc/4.0/를 참조하세요.
https://doi.org/10.1136/jitc-2021-004147
Statistics from Altmetric.com
68 readers on Mendeley
Request Permissions
If you wish to reuse any or all of this article please use the link below which will take you to the Copyright Clearance Center’s RightsLink service. You will be able to get a quick price and instant permission to reuse the content in many different ways.
Introduction
In recent years, the influence of the gut microbiome on remote organs, and mucosal and immune functions has been uncovered and exploited in the management of inflammatory conditions.1 Indeed, a role for the gut microbiome has been recognized in gastrointestinal disorders, and in the development and potential treatment of graft vs host disease, cardiovascular and neurodegenerative diseases, and cancer.2–5 Because of the possibility to influence the microbiome composition and the seemingly beneficial impact of this strategy on several disease states, research efforts have been focused on the identification of critical microbial-derived metabolites, and their therapeutic exploitation. In particular, short chain fatty acids (SCFAs) have recently gained specific attention for their impact on colon health, and for their immune-modulatory activities, which include both anti-inflammatory and tumor-suppressive functions.6 As their relative concentration can be sensitive to diet,7 and to prebiotics (complex carbohydrates that can be fermented by colonic bacteria) and probiotics (live bacteria that promote colonic health), increasingly being used in clinical practice,8 their modulation is raising interest as a therapeutic option.
소개
최근 몇 년간 장 미생물군이 원격 장기, 점막 및 면역 기능에 미치는 영향이 밝혀지고 염증성 질환 관리에 활용되고 있습니다.1 실제로 장 미생물군은 위장관 장애, 이식 대 호스트 질환의 발생 및 잠재적 치료, 심혈관 및 신경퇴행성 질환, 암 등에서 역할을 인정받고 있습니다.2–5 미생물군집 구성에 영향을 미칠 수 있는 가능성과 이 전략이 여러 질환 상태에 미치는 유익한 영향으로 인해, 연구 노력은 중요한 미생물 유래 대사물의 식별과 그 치료적 활용에 집중되어 왔습니다.
특히, 단쇄 지방산(SCFAs)은
대장 건강에 미치는 영향과 면역 조절 활동(항염증 및 종양 억제 기능 포함)으로 인해
최근 특별한 관심을 받고 있습니다.6
SCFAs의 상대적 농도는 식이 요인에 민감하며,7
장내 세균에 의해 발효될 수 있는 복합 탄수화물인 프리바이오틱스와 장 건강을 촉진하는 살아있는 세균인
프로바이오틱스에 영향을 받습니다.
이러한
프리바이오틱스와 프로바이오틱스는 임상实践에서 점점 더 널리 사용되고 있으며,8
SCFAs의 조절은 치료 옵션으로 관심을 끌고 있습니다.
SCFAs are part of the larger fatty acids (FAs) family, classified according to the length in carbon atoms, their degree of saturation (saturated, monounsaturated, and polyunsaturated) and their cisorientation or transorientation of double bonds (for a more extensive description of the general structure of FAs and their impact on lipid metabolism in T cells, please refer to Howie et al)9 SCFAs are saturated monocarboxylic FAs with up to five carbons in a single chain. These include formate (C1), acetate (C2), propionate (C3), butyrate (C4), and valerate (C5).10 They are distinguished from longer FAs, which include medium-chain (6–12 carbons, MCFAs), long-chain FAs (13–21 carbons, LCFAs), and very long-chain FAs (beyond 21) (a schematic representation of the most studied C2-C4, and examples of MCFAs and LCFAs are depicted in figure 1).
SCFAs는 지방산(FAs)의 큰 분류군에 속하며,
탄소 원자의 수, 포화도(포화, 단일불포화, 다중불포화) 및 이중 결합의 시스 또는 트랜스 배열에 따라 분류됩니다
(지방산의 일반적인 구조와 T 세포의 지질 대사への 영향에 대한 자세한 설명은 Howie et al을 참조하십시오)9
SCFAs는
단일 사슬에 최대 5개의 탄소 원자를 가진 포화 단일 카르복시산 지방산입니다.
이에는
포르메이트(C1),
아세테이트(C2),
프로피오네이트(C3),
부티레이트(C4),
발레레이트(C5)가 포함됩니다.10
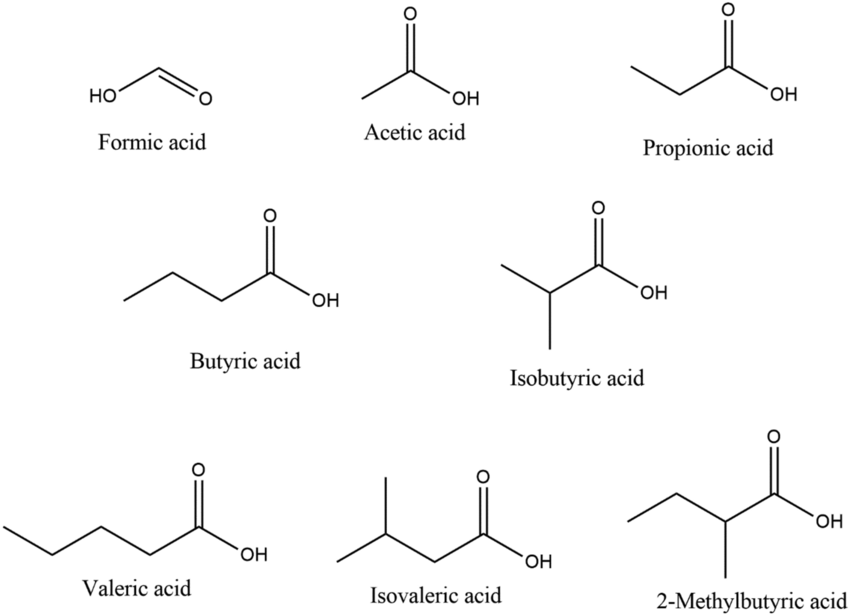
이들은
중간 사슬 지방산(6–12 탄소, MCFAs),
장사슬 지방산(13–21 탄소, LCFAs),
초장사슬 지방산(21 탄소 초과)과 구분됩니다
(가장 많이 연구된 C2-C4의 도식적 표현과 MCFAs 및 LCFAs의 예시는 figure 1에 표시되어 있습니다).
SCFAs are generated by gut microbes through the fermentation of non-digestible fibers and dietary carbohydrates, thus their concentration might vary according to the microbe composition and the anatomical location (refer to Cong et al for a recent comprehensive review).11 Thanks to shorter hydrophobic chains and the hydrophilic carboxyl group, SCFAs are water soluble and readily absorbed or transported into colon epithelial cells and used as the preferred energy substrates. In the proximal and distal portion of the human colon, the luminal concentrations of C2, C3, and C4 can, respectively, reach a relative concentration of ~130 nmol/Kg and 80 nmol/kg.12 13 In another study, Rombeau et al approximated SCFA concentrations in the content of the human colon to be 75 mM for acetate (C2), 30 mM for propionate (C3), and 20 mM for butyrate (C4).14 While a significant fraction is directly consumed by colonocytes, passive diffusion and active transport grant that micromolar concentrations are found in the portal blood and the liver (~250 µM for C2, 20–200 µM for C3, and 15–65 µM for C4), and also in the peripheral blood (20–150 µM for C2, 1–13 µM for C3, and 1–12 µM for C4),13 15 with various effects on different organ sites.16 The finding that all the major SCFAs are present in portal blood at concentrations several times greater than peripheral venous blood strongly suggests the colon is the major source of these FAs.17 18 Accordingly, dietary changes can alter their relative concentrations, as also infectious agents and health conditions.19 As an example, patients with cirrhosis revealed higher SCFA levels in the liver and in the portal vein than those measured in healthy controls.20
SCFAs는
장 내 미생물에 의해 소화되지 않는 식이 섬유와 탄수화물의 발효를 통해 생성되며,
따라서 미생물 구성과 해부학적 위치에 따라 농도가 달라질 수 있습니다(최근 종합적 검토는 Cong et al. 참조).11
짧은 친수성 사슬과 친수성 카르복실 그룹 덕분에 SCFAs는
수용성이며 대장 상피 세포로 쉽게 흡수되거나 운반되어 선호되는 에너지 기질로 사용됩니다.
인간 대장의 근위부와 원위부에서
C2, C3, C4의 장 내 농도는
각각 약 130 nmol/kg과 80 nmol/kg의 상대적 농도에 달할 수 있습니다.12 13
다른 연구에서 Rombeau 등(et al)은
인간 대장의 내용물에서 SCFA 농도를
아세테이트(C2) 75 mM, 프로피오네이트(C3) 30 mM, 부티레이트(C4) 20 mM로 추정했습니다.14
대장 상피세포에 의해 직접 소비되는 상당한 양에도 불구하고, 수동 확산과 능동 수송을 통해 미크로몰 농도가 문맥 혈액과 간 (~250 µM의 C2, 20–200 µM의 C3, 15–65 µM의 C4) 또한 말초 혈액에서도 검출됩니다(C2는 20–150 µM, C3는 1–13 µM, C4는 1–12 µM),13 15 이는 다양한 장기 부위에 다른 영향을 미칩니다.16 모든 주요 SCFA가 문맥 혈액에서 말초 정맥 혈액보다 수 배 높은 농도로 존재한다는 사실은 대장이 이러한 지방산의 주요 공급원임을 강력히 시사합니다.17 18
따라서
식이 변화는 그들의 상대적 농도를 변경할 수 있으며,
감염성 인자 및 건강 상태도 마찬가지입니다.19
예를 들어,
간경변 환자는 건강한 대조군보다 간과 문맥 정맥에서
더 높은 SCFA 수치를 나타냈습니다.20
Figure 1
Schematic representation of SCFA, MCFA and LCFA. The picture depicts the most common C2-C4 SCFAs, and representative members of the MCFA and LCFA family members.
T lymphocytes are sensitive and dependent on extra and intracellular FAs over the course of activation, proliferation, and memory differentiation. Numerous FA receptor/binding proteins regulate their relative representation in the extracellular environment, or their ability to signal via membrane-anchored or nuclear receptors.9 Several FAs act as ligands for nuclear receptors, a group of ligand-binding transcription factors and mediators of various metabolic and signaling pathways.21 In particular, long-chain polyunsaturated fatty acyls are considered the preferred ligands for the peroxisome proliferator-activated receptors,22 which are able to control lipid metabolism and Th1, Th2, Th17, and Treg cell differentiation.23 SCFAs instead are mostly known to act via G-protein-coupled receptor (GPCR) signaling, histone deacetylase (HDAC) inhibition, acetyl-CoA production, and metabolic integration.24 Multiple studies have established that T cell functions are sensitive to SCFA exposure, both in vitro and in vivo. Indeed, SCFAs can either promote a regulatory T cell phenotype or imprint T cells with effector functions, opening their possible exploitation to fine-tune adoptive T cell therapy (ACT) protocols. This review aims to highlight putative benefits and drawbacks of the use of SCFAs as immunomodulators, in both animal models and clinical studies, and to suggest how available knowledge can be translated to the most effective ACT protocols against cancer.
T 림프구는
활성화, 증식, 기억 분화 과정 전반에 걸쳐
세포 내외의 지방산(FAs)에 민감하고 의존적입니다.
수많은 지방산 수용체/결합 단백질은
세포 외 환경에서의 상대적 농도를 조절하거나,
세포막에 결합된 수용체나 핵 수용체를 통해 신호 전달 능력을 조절합니다.9
여러 지방산은 핵 수용체의 리간드로 작용하며,
이는 다양한 대사 및 신호 전달 경로의 조절자로 기능하는
리간드 결합 전사 인자 그룹입니다.21
특히 장쇄 다불포화 지방산 아실은
과산화체 증식 활성화 수용체(PPAR)의 선호되는 리간드로 간주되며,22
이는 지질 대사 및 Th1, Th2, Th17, 및 조절 T 세포 분화를 조절합니다.23
SCFA는 주
로 G-단백질 결합 수용체(GPCR) 신호전달,
히스톤 탈아세틸화 효소(HDAC) 억제,
아세틸-CoA 생산, 및 대사 통합을 통해 작용하는 것으로 알려져 있습니다.24
여러 연구에서
SCFA 노출이 체외 및 체내에서 T 세포 기능에 민감하다는 것이 입증되었습니다.
실제로 SCFAs는
조절성 T 세포 표현형을 촉진하거나
T 세포에 효과기 기능을 부여하여,
적응성 T 세포 치료(ACT) 프로토콜을 미세 조정하는 데 활용될 수 있는 가능성을 열어줍니다.
이 리뷰는 동물 모델과 임상 연구에서
SCFAs를 면역 조절제로 사용하기 위한 잠재적 이점과 한계점을 강조하고,
기존 지식을 암 치료를 위한 가장 효과적인 ACT 프로토콜로 전환하는 방법을 제안합니다.
SCFAs: mechanisms and specificities in T cells
SCFAs reach the cytoplasm either via passive diffusion across the plasma membrane or membrane transporters including MCT1 (monocarboxylate transporter-1/Slc16a1) and SMCT1 (sodium-coupled monocarboxylate transporter-1/Slc5 a8) or (summarized in figure 2A).25 MCT1 is a proton-linked monocarboxylic acid transporter, and a member of the MCTs family. MCT1 is constitutively expressed by colonic epithelial cells and upregulated over the course of T cell activation.26 It is well known for its role in transporting lactate, the most common short chain hydroxy- FA, which can be converted to other SCFAs by a sub-group of lactate-fermenting bacterial species.27 Its inhibition hinders T lymphocyte glycolysis,28 while promoting responses to anti-PD1 therapy.26 MCT1 can bind to the immunoglobulin family member CD147 or extracellular matrix metalloproteinase inducer (Emmprin or Basigin), which is also expressed on activated T cells,29 30 and was found to identify subsets of memory T cells in rheumatoid arthritis patients and of highly suppressive subset T regulatory cells.31 32 SMCT1 is a Na+-coupled electrogenic transporter for SCFAs. It has a preferred affinity for butyrate, followed by propionate, lactate, and acetate. It is predominantly expressed by colonocytes, dendritic cells (DCs), kidney, retinal and brain cells, while absent in T cells.33
SCFAs: T 세포에서의 메커니즘과 특이성
SCFAs는
세포질로 이동하는 두 가지 경로를 통해 세포질에 도달합니다.
첫째, 세포막을 통해 수동 확산으로 이동하거나,
둘째, MCT1(monocarboxylate transporter-1/Slc16a1) 및
SMCT1(sodium-coupled monocarboxylate transporter-1/Slc5 a8)과 같은
세포막 운반체를 통해 이동합니다(그림 2A에 요약됨).25
MCT1은 프로톤 결합형 단일카르복실산 운반체로, MCTs 가족의 일원입니다.
MCT1은 대장 상피 세포에서 지속적으로 발현되며 T 세포 활성화 과정에서 발현이 증가합니다.26
이는 가장 일반적인 단쇄 히드록시 지방산(SCFA)인 젖산을 운반하는 역할로 잘 알려져 있으며, 이 젖산은 젖산 발효 세균의 일부 하위 그룹에 의해 다른 SCFA로 전환될 수 있습니다.27 MCT1의 억제는 T 림프구의 글리코lysis를 방해하며,28 동시에 항-PD1 치료에 대한 반응을 촉진합니다.26 MCT1은 면역글로불린 가족 구성원인 CD147 또는 세포외 기질 금속 프로테아제 유도체(Emmprin 또는 Basigin)와 결합할 수 있으며, 이는 활성화된 T 세포에서도 발현됩니다.29 30 류마티스 관절염 환자의 기억 T 세포 하위 집합과 고도로 억제적인 T 조절 세포 하위 집합을 식별하는 데 사용되었습니다.31 32 SMCT1은 SCFAs를 운반하는 Na+-결합 전기적 운반체입니다. 이 단백질은 부티레이트에 대한 선호 친화성을 보이며, 그 다음으로 프로피오네이트, 락테이트, 아세테이트 순입니다. 주로 대장 상피세포, ден드리틱 세포(DCs), 신장, 망막 및 뇌 세포에서 발현되며, T 세포에서는 발현되지 않습니다.33
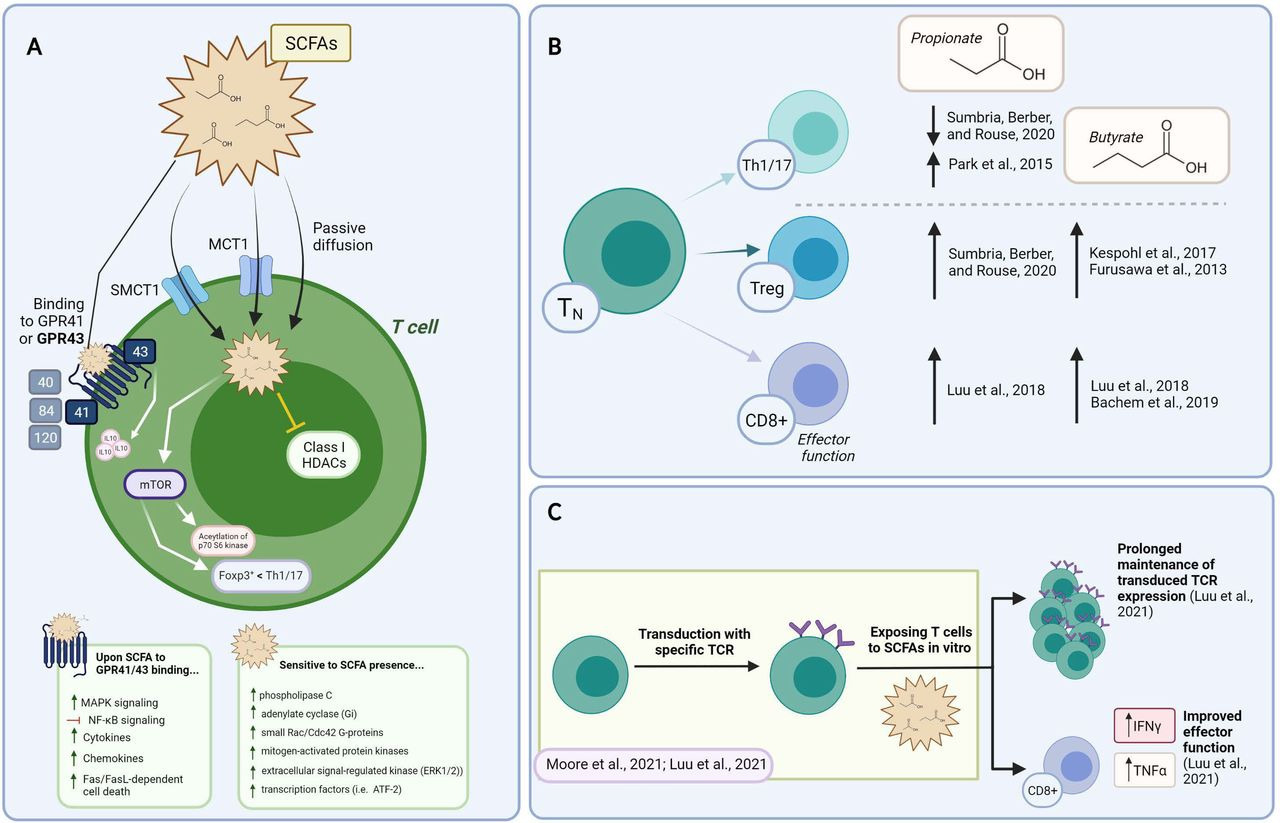




Figure 2
Constructing the mechanism of action and the impact of SCFAs on T cell functions in vitro. (A) Summary of SCFAs mechanism of action in T cells. The ability of SCFA to freely diffuse across the plasma membrane, be transported or bind to GPCR is shown. Their reported mechanism of action mediated by intracellular signaling events and HDAC is also depicted. (B) Supplementing T cell cultures with SCFAs impacts on Th1/17, Treg, and CD8+ T cell functional phenotypes (described within the text). Supporting references are indicated. (C) Exploiting SCFA over the course of TCR/CAR-T manufacturing. Data supports the ability of SCFAs to prolong transgene expression and increase effector functions. CAR, chimeric antigen receptor; HADS, histone deacetylase; SCFAs, short-chain fatty acids;
그림 2
SCFAs가 체외에서 T 세포 기능에 미치는 작용 메커니즘 및 영향.
(A) SCFAs의 T 세포 내 작용 메커니즘 요약. SCFAs가 세포막을 자유롭게 확산되거나 GPCR에 결합하거나 운반되는 능력이 표시되어 있습니다. 세포 내 신호 전달 사건과 HDAC를 통해 매개되는 보고된 작용 메커니즘도 설명되어 있습니다.
(B) T 세포 배양에 SCFAs를 보충하는 것이 Th1/17, Treg 및 CD8+ T 세포의 기능적 표현형에 미치는 영향 (본문 내에서 설명됨). 참고 문헌이 표시되어 있습니다.
(C) TCR/CAR-T 제조 과정에서 SCFA 활용. 데이터는 SCFA가 전유전자 발현을 연장하고 효과기 기능을 증가시킨다는 것을 지원합니다. CAR, 키메라 항원 수용체; HADS, 히스톤 탈아세틸화 효소; SCFAs, 단쇄 지방산;
SCFAs can also bind GPCRs on the cell membrane. Although the precise mechanism of action remains to be defined, SCFAs are thought to exert their functions mainly through GPCR-induced signaling, the inhibition of HDACs, metabolism tuning, and protein acetylation (summarized in figure 2A).34 Five GPRs with different binding affinity for FAs of various lengths have been described: GPR 40, 41, 43, 84, and 120. SCFAs mostly interact with GPR41 and 43, with different affinities for C2, C3 and C4, while GPR84 binds MCFAs, and GPR40 and GPR120 LCFAs (summarized in figure 2A).9 Another SCFA receptor belonging to a sub-class of GPCRs, the olfactory receptor 78 (Olfr78), senses C2 and C3. Most mature T cells have minimal expression of SCFA-sensing GPRs, except some memory CD8+ T cells which express Olfr78, and colonic regulatory T cells (Tregs) described to express GPR43.35 36 In addition, GPR43 has also been found expressed on colonic group 3 innate lymphoid cells (ILC s3) and on γδ T cells.37 Phospholipase C, adenylate cyclase, small Rac/Cdc42 G-proteins, mitogen-activated protein kinases (p38, c-Jun N-terminal kinase (JNK), extracellular signal-regulated kinase (ERK1/2)) and transcription factors (ie, ATF-2) are among the signaling molecules sensitive to SCFAs (summarized in figure 2A).38 Concentrations above 3 mM were reported to induce Fas upregulation and cause Fas/FasL-dependent cell death (summarized in figure 2A).39
SCFAs는 세포막상의 GPCR에 결합할 수 있습니다.
정확한 작용 메커니즘은 아직 명확히 규명되지 않았지만,
SCFAs는
주로 GPCR 유도 신호전달,
HDAC 억제,
대사 조절,
단백질 아세틸화 등을 통해 기능을 발휘하는 것으로 추정됩니다(그림 2A에 요약됨). 34
다양한 길이의 지방산에 대한 결합 친화성이 다른 5개의 GPR이 보고되었습니다: GPR 40, 41, 43, 84, 및 120. SCFAs는 주로 GPR41과 43과 상호작용하며, C2, C3, C4에 대한 결합 친화도가 다릅니다. 반면 GPR84는 MCFAs에 결합하며, GPR40과 GPR120은 LCFAs에 결합합니다(그림 2A에 요약됨).9 GPCR의 하위 클래스에 속하는 또 다른 SCFA 수용체인 후각 수용체 78(Olfr78)은 C2와 C3를 감지합니다.
성숙한 T 세포의 대부분은 SCFA 감지 GPR의 발현이 미미하지만, 일부 기억 CD8+ T 세포는 Olfr78을 발현하며, 대장 조절 T 세포(Tregs)는 GPR43을 발현하는 것으로 알려져 있습니다.35 36 또한 GPR43은 대장 그룹 3 선천성 림프구 세포(ILC s3)와 γδ T 세포에서도 발현되는 것으로 확인되었습니다.37 인산리파아제 C, 아데닐레이트 사이클레이즈, 소형 Rac/Cdc42 G단백질, 미토겐 활성화 단백질 키나제(p38, c-Jun N-말단 키나제(JNK), 세포외 신호 조절 키나제(ERK1/2)) 및 전사 인자(예: ATF-2)는 SCFA에 민감한 신호 분자 중 일부입니다. (그림 2A에 요약됨).38 3 mM 이상의 농도는 Fas 발현 증가를 유도하고 Fas/FasL 의존적 세포 사멸을 유발한다고 보고되었습니다(그림 2A에 요약됨).39
Data support the ability of SCFAs to mediate intracellular signaling events linked to the shaping of T cell fate, via both GPCR-dependent and independent mechanisms. For instance, Sun et al found that acetate (C2, 10 mM), propionate (C3, 0.5 mM), and butyrate (C4, 0.5 mM) favored the upregulation of IL-10 in Th1 cells with regulatory functions via GPR43 and mTOR and STAT3-dependent Blimp-1 expression (summarized in figure 2A).36 These authors also found that oral feeding of mice with butyrate (C4, 200 mM) in the drinking water recapitulated these events and protected mice from DSS-induced colitis.36 Likewise, Trompette et al found that GPR41 was needed for butyrate to improve cytotoxic T lymphocytes (CTL) expansion and effector functions.40 Indeed, provision of butyrate over the course of naïve CD8+ T cell activation in vitro improved the acquisition of cytolytic activities (0.5–1 mM), while oral administration of butyrate over a 2 week period (500 mM) boosted anti-viral T cell immunity.40 Pentanoate and butyrate (0.25–10 mM) were also recently described to augment IFNγ and TNFα secretion by activated CTLs via GPR41 and GPR43-independent mechanisms.35 41 In the case of γδ T cells, Dupraz et al found that propionate (10 mM) inhibited IL-17 and IL-22 production by intestinal murine and human γδ T cells, by a mechanism independent of GPR43 and also of MCT1.42 Thus, data generated so far suggests that SCFA play a role in lymphocyte differentiation, only in part through GPR41 and GPR43.
데이터는 SCFAs가 GPCR 의존적 및 비의존적 메커니즘을 통해 T 세포 운명 형성과 관련된 세포 내 신호전달 사건을 매개할 수 있음을 보여줍니다. 예를 들어, Sun 등(Sun et al)은 아세테이트(C2, 10 mM), 프로피오네이트(C3, 0.5 mM), 부티레이트(C4, 0.5 mM)가 GPR43 및 mTOR와 STAT3 의존적 Blimp-1 발현을 통해 조절 기능을 가진 Th1 세포에서 IL-10의 발현을 촉진한다는 것을 발견했습니다(그림 2A에 요약됨).36
이 연구자들은 또한 마우스에게 음용수에 부티레이트(C4, 200 mM)를 경구 투여했을 때 이러한 현상이 재현되었으며, DSS로 유발된 대장염으로부터 마우스를 보호한다는 사실을 발견했습니다.36 마찬가지로, Trompette 등 연구진은 GPR41이 부티레이트가 세포독성 T 림프구(CTL)의 증식과 효과기 기능을 개선하는 데 필요함을 발견했습니다.40 실제로, 체외에서 미분화 CD8+ T 세포 활성화 과정에서 부티레이트를 공급하면 세포독성 활성 획득이 개선되었습니다(0.5–1 mM), 반면 2주 동안 경구 투여한 부티레이트 (500 mM)는 항바이러스 T 세포 면역력을 강화했습니다.40 펜타노산과 부티레이트(0.25–10 mM)는 최근 활성화된 CTL의 IFNγ 및 TNFα 분비를 GPR41 및 GPR43 독립적 메커니즘을 통해 증강시킨다는 것이 보고되었습니다.35 41 γδ T 세포의 경우, Dupraz 등 연구진은 프로피오네이트(10 mM)가 장 내 마우스 및 인간 γδ T 세포의 IL-17 및 IL-22 생산을 GPR43 및 MCT1과 독립적인 메커니즘을 통해 억제한다는 사실을 발견했습니다.42 따라서 현재까지 생성된 데이터는 SCFA가 림프구 분화에 역할을 하며, 이는 부분적으로 GPR41 및 GPR43을 통해 이루어진다는 것을 시사합니다.
Other important functions of SCFAs reside in their role as a source of acetyl-CoA and also in their ability to inhibit HDACs. Indeed, SCFAs can be converted to acetyl-CoA, which fuels major metabolic processes, such as the mitochondrial tricarboxylic acid (TCA) cycle, FA synthesis, and protein acetylation. When integrated into the Krebs cycle, acetyl-CoA increases energy (ATP/ADP) production, leading to mTOR activation. This has been found to favor T cell differentiation into effector T cells such as Th1 and Th17 cells at the expense of FoxP3+ T cells (summarized in figure 2A).25 Acetyl groups derived from acetyl-CoA are also used by histone acetyltransferases (HATs) to decorate histone tails and promote gene transcription.43 This has been described in epithelial cells where SCFAs were found to regulate HAT and hypoxia inducible factor (HIF) stability.44 Also, non-histone proteins, including signaling molecules and transcription factors, can undergo acetylation in response to SCFA, ultimately regulating transcriptional processes.45 For instance, in CD4+ T cells activated in Th17 conditions, Park et al reported that acetate (10 mM), propionate (1 mM) and butyrate (0.1–0.5 mM) controlled acetylation of p70 S6 kinase, downstream to mTOR signaling independently of GPR41 and GPR43 (summarized in figure 2A).25 Accordingly, propionate and butyrate were found to be capable of inhibiting class I HDACs.46 47 These remove acetyl groups, leading to the tightening of chromatin and transcriptional repression. In T cells, the FoxP3 and IL-10 gene loci are targets of such regulation.48 Via their activities as HDAC inhibitors, acetate, propionate and butyrate also enhance aryl hydrocarbon receptor (AhR) ligand-induced responses in gene- and cell context-dependent events.20 49 50 Although whether SCFAs control HDAC activity directly or indirectly requires further investigation, it is interesting to note that these metabolites can simultaneously increase availability of acetyl groups and block HDACs, jointly increasing acetylation events and favoring gene expression. Additional studies will improve our understanding of SCFA-controlled events linked to metabolic and epigenetic pathways.
SCFA의 다른 중요한 기능은 아세틸-CoA의 공급원으로서의 역할과 HDAC 억제 능력에 있습니다. 실제로 SCFAs는 아세틸-CoA로 전환되어 미토콘드리아 트리카르복실산(TCA) 회로, 지방산 합성, 단백질 아세틸화 등 주요 대사 과정에 에너지를 공급합니다. 아세틸-CoA가 크렙스 회로에 통합되면 에너지(ATP/ADP) 생산이 증가하여 mTOR 활성화를 유도합니다. 이는 FoxP3+ T 세포 대신 Th1 및 Th17 세포와 같은 효과기 T 세포로의 분화를 촉진하는 것으로 확인되었습니다(그림 2A에 요약됨).25 아세틸-CoA에서 유래한 아세틸 그룹은 히스톤 아세틸전달효소(HATs)에 의해 히스톤 꼬리를 장식하고 유전자 전사를 촉진하는 데 사용됩니다.43 이는 상피 세포에서 SCFA가 HAT 및 저산소 유도 인자(HIF)의 안정성을 조절한다는 점에서 설명되었습니다.44 또한 히스톤이 아닌 단백질, 예를 들어 신호 전달 분자 및 전사 인자도 SCFA에 반응하여 아세틸화될 수 있으며, 이는 최종적으로 전사 과정을 조절합니다.45 예를 들어, Th17 조건에서 활성화된 CD4+ T 세포에서 Park 등 연구진은 아세테이트(10 mM), 프로피오네이트(1 mM), 부티레이트(0.1–0.5 mM)가 GPR41 및 GPR43에 독립적으로 mTOR 신호전달 하류의 p70 S6 키나제의 아세틸화를 조절한다는 것을 보고했습니다(그림 2A에 요약됨).25 이에 따라 프로피오네이트와 부티레이트는 클래스 I HDAC를 억제하는 것으로 확인되었습니다.46 47 이 HDAC는 아세틸 그룹을 제거하여 염색질의 조밀도를 높이고 전사 억제를 유발합니다. T 세포에서 FoxP3 및 IL-10 유전자 위치는 이러한 조절의 대상입니다.48 HDAC 억제제로서의 활성을 통해 아세테이트, 프로피오네이트 및 부티레이트는 유전자 및 세포 맥락에 따라 아릴 하이드로카본 수용체(AhR) 리간드 유도 반응을 강화합니다.20 49 50 SCFAs가 HDAC 활성을 직접적으로 또는 간접적으로 조절하는지는 추가 연구가 필요하지만, 이러한 대사산물이 아세틸 그룹의 가용성을 증가시키고 HDAC를 차단하여 아세틸화 사건을 증가시키고 유전자 발현을 촉진한다는 점은 흥미로운 점입니다. 추가 연구는 SCFA에 의해 조절되는 대사 및 에피게노믹 경로와 관련된 사건에 대한 이해를 높일 것입니다.
SCFAs shape T cell differentiation in vitro
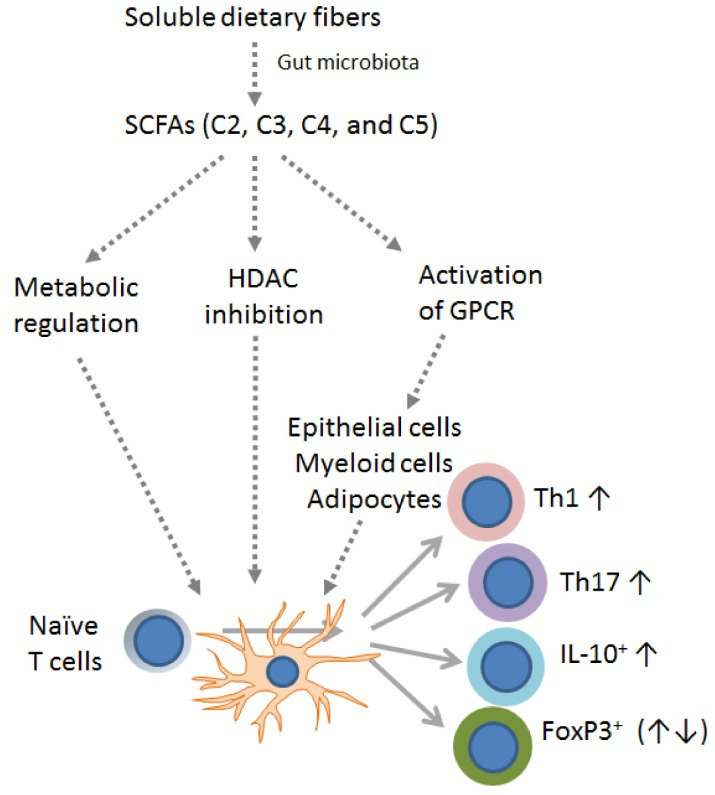
Antigen recognition via the T cell receptor (TCR) and the engagement of costimulatory and cytokine receptors induces a complex signaling cascade that leads to the activation, proliferation, and differentiation of naïve T cells and the acquisition of effector or regulatory activities. Effector T cells fight pathogens and cause tissue inflammation, while regulatory T cells counterbalance such events. T cell differentiation involves heritable changes to the epigenetic landscape, and also metabolic rewiring involving Akt-mTOR (mammalian target of rapamycin) signaling.51 52 While naïve T cells mostly rely on the TCA cycle and oxidative phosphorylation, activated T cells switch to glycolysis, glutaminolysis and FA synthesis to accommodate an increased demand for energy.53 Nutrients and metabolites provide significant regulatory signals for metabolic adaptation and ensuing T cell differentiation. It is now clear that the gut microbiota as well as SCFAs, in the form of acetate, butyrate, and propionate shape the generation of both effector and regulatory T cells via epigenetic and metabolic events.54
In the case of CD4+ T cells, the effects of SCFAs vary according to the activation conditions and the cytokine milieu. Furusawa et al found that the addition of 0.1 mM butyrate to cultures of CD4+ T cells activated with anti-CD3 and -CD28 antibodies, TGF-β1 and IL-2, promoted an increase in genome-wide histone H3 acetylation and Foxp3 expression, leading to Treg differentiation (summarized in figure 2B).55 More recently, Kespohl et al extended this study and found that while low butyrate concentrations (0.1–0.5 mM) facilitated the differentiation of Foxp3+ Tregs, higher concentrations (1 mM) induced the expression of T-bet and IFNγ via the control of promoters histone acetylation (summarized in figure 2B).56 Park et al also found that titrating amounts of acetate (1–10 mM) and propionate (0.1–1 mM) and butyrate (0-1-0.5mM) supported Th1 or Th17 effector cell differentiation via HDAC inhibition and independently of GPR41or GPR43. These SCFA-conditioned cells, however, also produced IL-10 proving less inflammatory in vivo compared unconditioned ones (summarized in figure 2B).25 In another study, increasing concentrations of propionate (100–400 uM) in 5-day CD3/CD28 cultures of naïve CD4+ T cells inhibited Th1 and Th17 T cells differentiation, while promoting FoxP3+ Tregs (summarized in figure 2B).57 Yang et al also found that SCFAs (acetate, 10 mM; propionate, 0.5 mM; butyrate, 0.5 mM) promoted IL-22 production in CD4+ T cells and ILCs through GPR41 and HDAC inhibition and the expression of the AhR and the HIF1α transcription factors.50 While results might appear contradictory, it should be noted that the level of CD3 stimulation, the cytokine milieu, and the relative concentration of SCFAs adopted in the above-mentioned studies varied, suggesting that SCFAs might balance CD4+ T cell regulatory/effector function according to the milieu. In the case of CD8+ T cells, exposure to 1 mM propionate and butyrate was mostly shown to augment expression of IFNγ and Granzyme B (GzB), and effector function at large (summarized in figure 2B).41 Mechanistically, butyrate increased acetylation of histone H4 at Tbx21 and IFNγ promoters, supporting an HDAC-dependent mechanism. Of note in this study, acetate also promoted IFNγ expression, but via a mechanism less dependent on HDAC inhibition, and rather linked to CTLs metabolism. This observation is in line with a previous report showing that acetate conversion to acetyl-CoA supported GAPDH/glycolytic activity, linked to rapid recall responses of memory CD8+ T cells.58 Similarly, Qiu et al reported that 1 mM acetate could rescue IFNγ expression by CD8+ T cells cultured in glucose-restricted conditions, by controlling histone acetylation and chromatin accessibility in an acetyl-CoA synthetase-dependent manner. The authors found that the same applied to exhausted T cells, through which the ability to express IFNγ was rescued by ex vivo acetate supplementation.59 Bachem et al more recently found that SCFAs also promote long-lasting memory phenotypes.60 In this work, authors found that acetate, propionate, and butyrate (0.5 mM) were able to support long-term survival of antigen-activated T cells, through the upregulation of FoxO1, a transcription factor required for memory T cell differentiation. These authors also reported that the transition to a memory phenotype was due to butyrate uncoupling the TCA from glycolysis, favoring the use of FA and glutaminolysis (summarized in figure 2B).60
Thus, there is evidence supporting the ability of SCFAs to influence T cell differentiation in vitro, via cell-autonomous events involving epigenetic and/or metabolic regulation. Although these effects might be peculiar to supraphysiological concentrations (in the mM range, ie, ~1000 and ~100 fold higher than those recently measured in the peripheral and the hepatic portal blood61) data also support the notion that SCFAs are active in vivo. Indeed, they can promote tolerogenic and anti-inflammatory profiles (by skewing Treg phenotypes), and also effector and memory functions (Th1/Th17, CTL), possibly via the integration of extracellular clues (TCR/CD28 stimuli, cytokines, nutrients). Such contrasting effects might account for some discrepant reports in vivo, and yet represent solid ground for SCFA exploitation in ACT protocols (summarized in figure 2C).
SCFAs have immunomodulatory effects on T cell-dependent responses in vivo
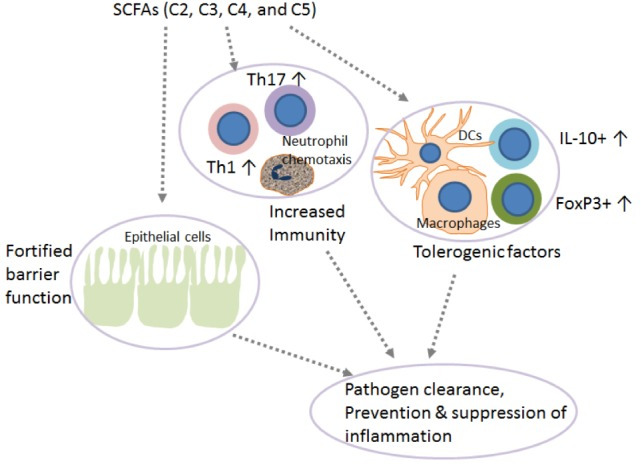
SCFAs in vivo have been reported to induce IL-10–expressing Foxp3+ and Foxp3− regulatory T cells and antagonize inflammation or effector T cells such as Th1 and Th17 cells and mediate inflammatory responses, depending on the physiological state, the immunological and the pathological milieu, and the relative SCFAs abundance. Beneficial anti-inflammatory functions of SCFAs were observed in several disease models, including colitis, and colitis-associated colon cancer, autoimmune manifestations, airway disease, metabolic syndrome, and ischemia-reperfusion injury of the kidney.6 62 In vivo, acetate, propionate, and butyrate feeding, was also shown to dampen immune responses via regulatory T cells,35 48 and also DCs,17 63 macrophages17 64 and neutrophils65 (summarized in figure 3A). Such effects were demonstrated in models as small as embryonic zebrafish, where exposing wound sites to butyrate (30 mM) significantly reduced the recruitment of M1-type proinflammatory macrophages, ultimately revealing anti-inflammatory activity.66 In a mouse model of corneal infection, propionate feeding (500 mM over 3 weeks) was found to have immunomodulatory effects at ocular lesion sites by concomitantly increasing Treg representation and reducing Th1 and Th17 pro-inflammatory T cells (summarized in figure 3A).57 Likewise, oral administration of propionate (150–200 mM) augmented Treg suppressive activity in an experimental autoimmune encephalomyelitis model,67 and Treg expansion in hypertensive cardiovascular damage models.68 Propionate was also reported to lower the number of proinflammatory Th17 cells in these models,68 and to hinder IL-17 production by mouse and human intestinal γδ T cells.42 Additionally, in a preclinical non-obese diabetic (NOD) model for autoimmune diabetes, acetate supplementation (200 mM) reduced the disease incidence, while butyrate or acetate-fortified diets respectively increased Treg number and activity and reduced autoreactive CD8+ effector T cells in lymphoid tissues.61 Nevertheless, supraphysiological doses of orally administered SCFAs (200 mM over 4 weeks) induced Th1 and Th17 effectors and IL-10+ regulatory T cells in ureter and kidney tissues, overall leading to T cell-mediated ureteritis, and kidney hydronephrosis.69 In two independent studies, administration of butyrate or a mix of acetate, butyrate, and propionate revealed protective effects against the development and the growth of colorectal cancer.70 71 Whether protective effects involved changes in T cell functions was not investigated.
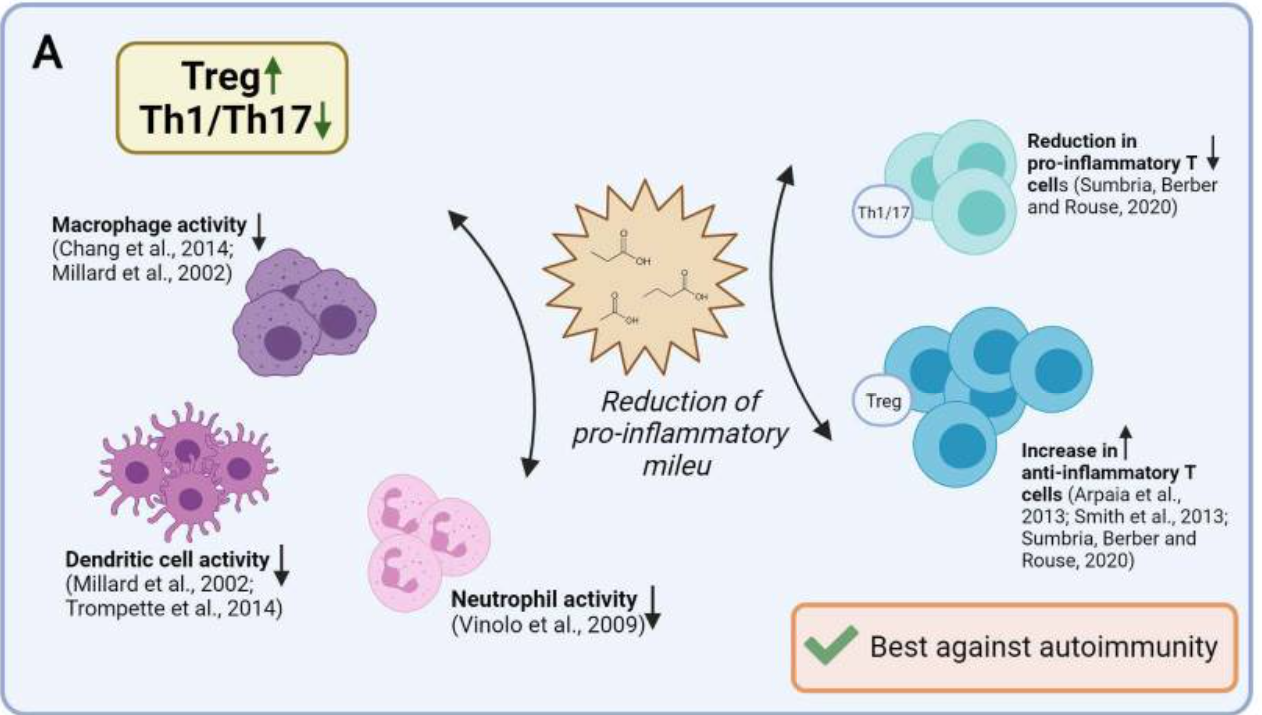

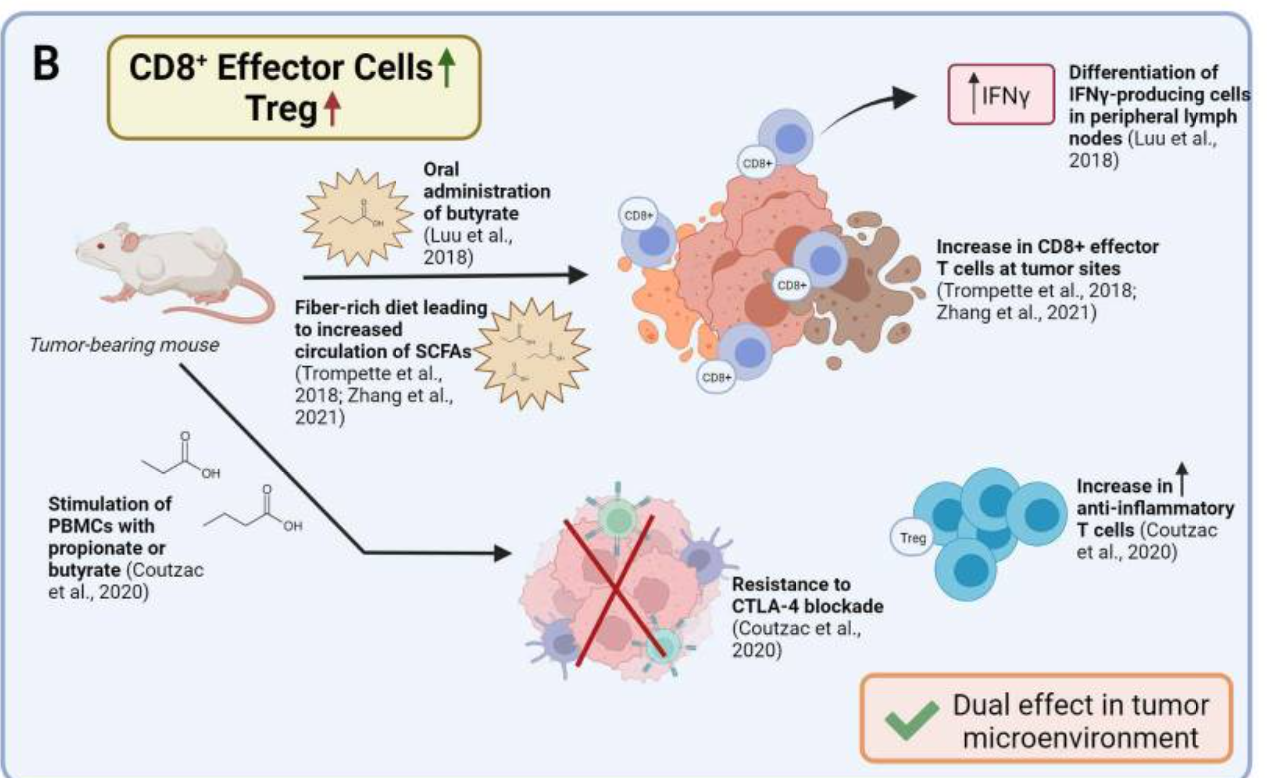

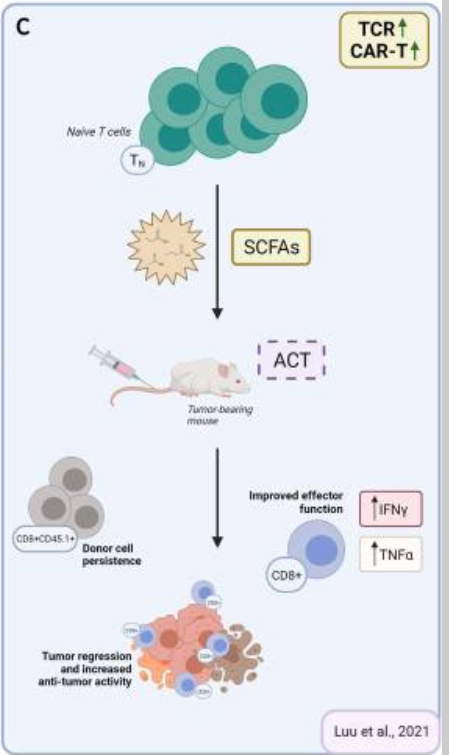

Figure 3
Reported effects of SCFAs on T cell functions in vivo and possible application for ACT strategies. (A) Reported effects of SCFAs in autoimmune manifestations. SCFAs can dampen the pro-inflammatory activities of macrophages, dendritic cells, and neutrophils, and Th1/17 cells, while upregulating anti-inflammatory Treg cells. (B) Reported pro- and anti-inflammatory activities of SCFAs in the context of cancer. Studies in tumor-bearing mice demonstrate that in vivo administration of SCFAs or of fiber rich diets can promote CD8+ T cell effector functions, but also increase the representation of Tregs. This is evidence for SCFAs having counter-regulatory effects according to the disease or the therapeutic setting. (C) Suggested use of SCFA in ACT settings. To take advantage of SCFAs in the context of ACT and overcome possibly pleiotropic effects observed by in vivo administration, preconditioning of T cell product by SCFAs supplementation represents a valuable strategy. ACT, adoptive T cell therapy; SCFAs, short-chain fatty acids.
Results from human trials reported so far appeared to support an anti-inflammatory effect secondary to SCFA administration to patients. SCFA supplementation was indeed tested in patients with multiple sclerosis patients, due to their reduced propionate concentration compared with healthy controls. Daily dietary supplementation with propionate was reported to restore plasma propionate concentration and also immunological parameters in MS patients with MS to those of healthy individuals.72 When tested as an adjunct to disease modifying therapy, propionate promoted an increase in Treg cell numbers and function, and a significant decrease in Th1 and Th17 cells. This was paralleled by a reduced annual relapse rate and delayed disease progression, although participant numbers were low and results should be considered with caution.73 In another trial, a propionate prescription to end-stage renal disease human patients significantly reduced C reactive protein and this correlated with the expansion of Treg cells in circulation.74 In contrast, a randomized, controlled type 1 diabetes clinical trial recently failed to report significant changes to CD4+ and CD8+ T cells, B cells, and natural killer cells in circulation after oral butyrate administration.75 The authors suggested the contrasting data observed in the clinical trial compared with previous preclinical studies could be due to inconsistencies in administration, dosing, and timing, which are all critical factors to consider when establishing a standardized protocol of SCFAs for in vivo administration.
In contrast to the above-mentioned studies, reports related to CD8+ T cell-driven immunity are in favor of SCFAs mostly having pro-inflammatory effects. Indeed, Luu et al found that exposing CD8+ T cells to butyrate in vivo ameliorated CTL function and persistence. While pretreating Ly5.1+ CD8+ T cells with 1 mM butyrate for 3 days imprinted T cells for long-lasting effector in vitro functions, oral administration of butyrate promoted differentiation of IFNγ-producing cells in peripheral lymph nodes (summarized in figure 3B).41 These effects were found independently of T cells expressing GPR41 and GPR43 and relied instead on butyrate-induced HDAC inhibition and metabolic reprogramming. In line with these findings, the administration of SCFA-rich, high fiber diets was associated with augmented CD8+ effector T cell function at sites of inflammation or tumors (summarized in figure 3B).40 76 In the case of infection, the exposure of mice to high fiber diets blunted CXCL1 production and neutrophil recruitment to infected lungs, limiting tissue immunopathology, while promoting metabolic activity and effector function of CD8+ T cells.40 Likewise, in tumor-bearing mice, administration of pectin, a fiber found in vegetable and fruits, improved CD8 effector T cell responses, and sensitivity to anti-PD-1 therapy in a mouse model of colon carcinoma.76 Dose-dependent effects might explain discrepancies, which should be further investigated in models, allowing to dissect the relative contribution of CD4 and CD8 regulatory/effector subsets. The possibility to reach relative subset balance via controlled diets and ad hoc pre/post-biotic supplementation yet appears a pursuable approach.
A role of SCFAs in chemotherapy, radiotherapy and immunotherapy of cancer
Preclinical and clinical reports indicate that chemotherapy or radiotherapy induces major changes in the composition of the gut microbiota. Data also indicate that anticancer therapeutic efficacy of chemotherapyor radiotherapy, and also of immunotherapy benefits of selected gut microbiota composition. This is because commensal microbes, and their SCFA derivatives (as well as antibiotic treatments) directly modulate cancer development and growth, tune intestinal inflammation, and influence efficacy and safety of anticancer therapy.70 This is most evident for, but not limited to, colon cancer. Indeed, SCFAs can boost barrier immunity, limiting persistent bacterial invasion secondary to the damaged gut barrier, the latter of which is responsible for chronic inflammatory responses and colon cancer progression. Concomitantly, SCFAs can also mediate therapy resistance.77
The impact of the gut microbiome on chemotherapy efficacy and toxicity has long been recognized.78 Chemotherapy (and also radiotherapy) often cause intestinal microbial dysbiosis, which leads to intestinal mucositis due to a number of effects: (1) inflammation and oxidative stress, (2) gastrointestinal permeability destruction, (3) mucus layer formation alteration, (4) epithelial repair, and (5) secretion of immune factors.70 Provision of SCFA-producing bacteria contributed to mucosa recovery and intestinal homeostasis. Direct application of SCFAs such as butyrate reported to increase IL-10 and reduce IL-12 and TNF-α in chemotherapy-induced colitis.79 Instead, recent work by He et al demonstrated that butyrate in combination with oxaliplatin promoted antitumor immunity. It favored CTL effector function in MC38 colorectal cancer tumor-bearing mice, ameliorating tumor control.80 Of note, in this setting, butyrate supplementation in vivo also granted better therapeutic activity to anti-PD-1 immune checkpoint blocker (ICB) therapy.80 This result is in line with a recent clinical study proving the efficacy of the association of SCFAs and PD-1 ICB in solid cancers.81
Radiotherapy efficacy and safety has also been reported to be influenced by the microbiome composition and SCFA representation. Radiation-induced intestinal injury involves a decrease in the diversity of intestinal flora and the concentration of SCFAs.82 In a recent report, Guo et al found that selected gut microbial families (Lachnospiraceae and Enterococcaceae) were associated with radioprotection both in patients and mice, and that treatment with propionate (and tryptophan metabolites) attenuated DNA damage and the release of reactive oxygen species in hematopoietic and gastrointestinal tissues, mitigating proinflammatory responses and promoting hematopoiesis and intestinal repair after radiation.83 Zhu et al found that non myeloablative total body irradiation, frequently used to precondition patients and create ‘space’ for adoptive T cell therapy products, impeded the growth and maintenance of SCFA-producing bacteria in the intestine with consequences on intestinal barrier integrity and functionality, ultimately impacting immune cell subset representation.84 A recent study by Yang et al found that the presence of butyrate and/or butyrate-producing bacteria hindered antitumor responses to irradiation.85 When mice were given a gram-positive-targeting antibiotic known to select for butyrate-producing bacteria prior to irradiation treatment, outcomes were improved, in both the MC38 colorectal cancer and B16 melanoma transplantable tumor models. Mechanistically, intratumoral accumulation of butyrate suppressed the activation of STING and production of IFNβ in DCs within the tumor, lowering protective T cell responses.85
In the case of immunotherapy, the microbiome composition has been reported to enhance or dampen antitumor responses, according to the therapeutic setting, and especially in the case of ICBs.86 In an experimental subcutaneous tumor model, a defined commensal consortium made of human bacterial strains elicited strong CD8+ T cell-mediated antitumor immunity, also improving responses to checkpoint blockers.87 In clinical settings, the enrichment of Faecalibacterium and other genera belonging to the phyla Firmicutes was similarly associated with beneficial clinical response to ICBs.88 Also the relative abundance of Akkermansia muciniphila was found a useful clinical predictor of poor responses to ICB in patients with lung and kidney cancers. This was paralleled by the observations that antibiotic-treatments correlated with lower, progression-free survival and overall survival when compared with untreated controls, both in patients and in mice, and that bacteria supplementation to antibiotic-treated mice restored the response to anti-PD-1 therapy.89 In a parallel study, it was found that efficacy of anti–PD-L1 therapy correlated with improved representation of Bifidobacterium longum, Collinsella aerofaciens, and Enterococcus faecium.90 In line with these studies, first-in-human clinical trials recently reported that fecal microbiota transplantation can affect how metastatic melanoma patients respond to anti-PD-1 immunotherapy. Results from these trials highlighted how the gut microbiome regulated changes in distinct proteomic and metabolomic signatures previously shown to be associated with responses to anti-PD-1, increased CD8+ T cell activation, and decreased frequency of interleukin-8-expressing myeloid cells.91
Nevertheless, in a recent study, Coutzac et al observed that, in mouse models of melanoma and in metastatic melanoma patients, high serum levels of butyrate and propionate were associated with resistance to CTLA-4 blockade and a higher proportion of Treg cells. Butyrate supplementation to melanoma tumor-bearing mice in combination with αCTLA-4 restrained DC maturation and the accumulation of both effector and memory tumor-specific CTLs. Likewise, when baseline PBMC samples from metastatic melanoma patients were stimulated with increasing concentration of butyrate ex vivo, there was a correspondent increase in Tregs (summarized in figure 3B).92 Thus, while SCFAs can promote potent CTLs, they can also favor Treg accumulation, and restrain DC maturation.
It should however be considered that while several reports emphasize a role for the microbiome and microbiome-derived SCFAs in vivo in promoting responses to therapy, data also underline the ability of these compounds to exert anti-inflammatory activities, ultimately supporting complex pleiotropic effects and thus, the difficulty in predicting clear outcomes. Indeed, timing of administration, relative abundance, and the physiological or disease context can simultaneously influence the consequence of SCFA exposure. We argue that these are important variables to consider when envisaging the exploitation of SCFAs in the setting of ACT.
SCFAs for the development of an effective ACT protocol
ACT foresees the in vitro expansion/generation of large numbers of tumor-specific T cells and infusion to patients. Tumor-reactive T cells can be taken from tumor samples and expanded into sufficient numbers, or from peripheral blood and genetically-engineered with TCR or chimeric antigen receptor (CAR).93–95 TCR-redirected T cells recognize both extracellular and intracellular antigens presented in the context of major histocompatibility complex molecules (MHC/HLA) molecules, while CAR-T cells bind surface antigens without MHC/HLA restrictions.96 T cell cloning, and TCR sequencing,97 and the isolation of several single-chain antibodies95 has allowed the isolation of a number of TCR and of synthetic CAR molecules with optimized antigen binding and signaling abilities. In addition, the identification of T cell subsets endowed with effector functions or memory potential, together with the exploitation of homeostatic cytokines, such as IL-7 and IL-15, has allowed to optimize manufacturing processes to grant extensive T cell expansion in the absence of terminal differentiation.98–100
It has been recognized that ideal T cell products should have potential for rapid memory recall responses, homing to appropriate anatomical locations, in vivo re-expansion and also long-term survival. Given the notion that SCFAs can fine-tune T cell responsiveness and memory formation, their use over the course of manufacturing has recently been explored. Vodnala et al studied the phenotype of tumor infiltrating lymphocytes (TILS) and found that conditions that determine a diminished uptake and consumption of local nutrients (such as necrosis that leads to a profound alteration of the potassium ion gradient within the tumor microenvironment) leads to a reduction in acetyl coenzyme A (AcCoA).101 This caused a decrease in histone acetylation and silencing of effector genes. They also proved that treating expanding T cells with acetate (5 mM), the immediate precursor of AcCoA, restored nucleo/cytosolic AcCoA and IFNγ production, reversing stemness programs and enabling the acquisition of effector programs. These data underline the notion that nutrient availability and processing controls metabolic features and acts to epigenetically imprint T cell fate.101 Moore et al investigated the impact of combining homeostatic cytokines and SCFAs over T cell manufacturing. They found that T cells expressing a tyrosinase-reactive TCR (TIL 1383I) gradually lost TCR expression over a 12-day long culture. HDAC inhibition reverted this event, suggesting that transgene downregulation observed during T cell cultures was epigenetically regulated. In this study, butyrate supplementation (1 mM) preserved the expression of the TIL 1383I TCR in both lenti-transduced and retroviral-transduced CD4+ and CD8+ T cells (summarized in figure 2C).102 SCFAs or HDAC inhibitors-imprinted long-lasting transgene expression correlated with improved therapeutic activities. Luu et al also investigated the use of SCFAs on antigen-specific CD8+ CTLs and CAR T cells in adoptive T cell therapy settings. Treatment with pentanoate and butyrate improved CTLs and CAR T cells effector functions, which correlated with increased antitumor reactivity and superior therapeutic outcomes in syngeneic murine melanoma and pancreatic cancer models (summarized in figures 2C and 3C).103 SCFA-reprogramming improved CD25, IFNγ and TNF-α expression, via GPR41/GPR43-independent events involving HDAC-inhibition and metabolic modulation. The authors also validated the bacterial source producing the highest amount of pentanoate and butyrate among the top 14 bacterial strains representing the expected microbiome profile of the human intestine: Megasphaera massiliensis of the phyla Actinobacteria. Supernatant derived from M. massiliensis had a significant impact on the production of IFNγ and TNFα of CD8+ effector T cells, demonstrating that the presence of specific microbial strains can have a direct influence over T cell function in vivo. In parallel studies, we similarly found that supplementing IL-7/IL-15 driven cultures of TCR transduced T cells with propionate (1–10 mM) or butyrate (0.2–1 mM) favored the expansion of IFNγ-producing and TNFα-producing cells, without causing their terminal differentiation (unpublished data). This represents an interesting phenotype given that such T cell products might simultaneously be capable of immediate effector function and also of long-term persistence.
Thus, although further studies would be needed to better understand SCFA-instructed phenotypes, including the epigenetic and metabolic events subtending the observed effects, current data support the use of SCFAs during T cell manufacturing. SCFAs imprint genetically engineered T cells with more stable TCR/CAR transgene expression, optimal functionality and memory potential, and in vivo improved therapeutic activity. While it might be tempting to suggest that supplementary SCFA treatments might further support engineered T cells on in vivo transfer, data available to-date might also predict counter-productive effects due to pleiotropic SCFA activities. Of note, searching the NIH clinicaltrial.gov site with ‘SCFA’ or ‘SCFA and immune’ as keywords identified 557 and 72 ongoing trials, respectively. Such a considerable number of trials likely reflects the fact that propionate is classified as a food product in the European Union and the USA, and thus generally considered safe, and as such of interest as a natural immune modulator. It is predictable that new data on the ability of SCFAs to shape the immune landscape within and outside the gut might thus become available soon, and better instructs their therapeutic exploitation in patients.
Conclusions
ACT with TILs or TCR/CAR-engineered T cells represents a valuable therapeutic opportunity to fight cancer. While highly efficacious in the setting of some hematological malignancies, optimization is still needed and required in the setting of solid tumors to improve efficacy while limiting toxicity. Strategies able to ameliorate functional and survival potential of T cell products are continuously being tested to improve efficacy and reduce treatment related toxicity and costs. Several factors can influence T cell engraftment and persistence. The microbiota and microbiota-derived components have been shown to contribute to the success of chemotherapy and radiotherapy and of ICB. SCFAs represent important links between the microbiota and the immune system. They have been shown to impact T cell function, and to epigenetically imprint T cell products with superior transgene expression, cytokine production, and survival. However, as the relative SCFA representation and local concentration at given anatomical districts remains largely unknown and given the SCFA pleiotropic effects on various immune and non-immune cell subsets (T cells, neutrophils, antigen presenting cells and epithelial cells) their direct exploitation in vivo might be currently questioned. Nevertheless, their adoption in vitro even at supraphysiological concentrations as a supplement in the generation of T cell product appears very approachable. Then, once the complex host–microbiota interplay would be more comprehensively understood, in vivo supplementation might be envisaged, and dosing, scheduling, and route of delivery optimized to balance local versus distal, proinflammatory versus anti-inflammatory effects, and ameliorate conventional and more innovative anticancer treatments.
Ethics statements
Patient consent for publication
Not applicable.
Acknowledgments
The authors wish to thank Dr Arianna Pocaterra and Dr Marco Catucci (IRCCS Ospedale San Raffaele, Milan, Italy) for critical reading of the manuscript. Figures were created with BioRender.com.
References
- ↵
- Durack J,
- Lynch SV
- ↵
- Cheng WY,
- Wu C-Y,
- Yu J
- ↵
- Mitra S,
- Drautz-Moses DI,
- Alhede M, et al
- ↵
- Ghaisas S,
- Maher J,
- Kanthasamy A
- ↵
- Mathewson ND,
- Jenq R,
- Mathew AV, et al
- ↵
- Hosseinkhani F,
- Heinken A,
- Thiele I, et al
- ↵
- Tan J,
- McKenzie C,
- Potamitis M, et al
- ↵
- Salminen S,
- Collado MC,
- Endo A, et al
- ↵
- Howie D,
- Ten Bokum A,
- Necula AS, et al