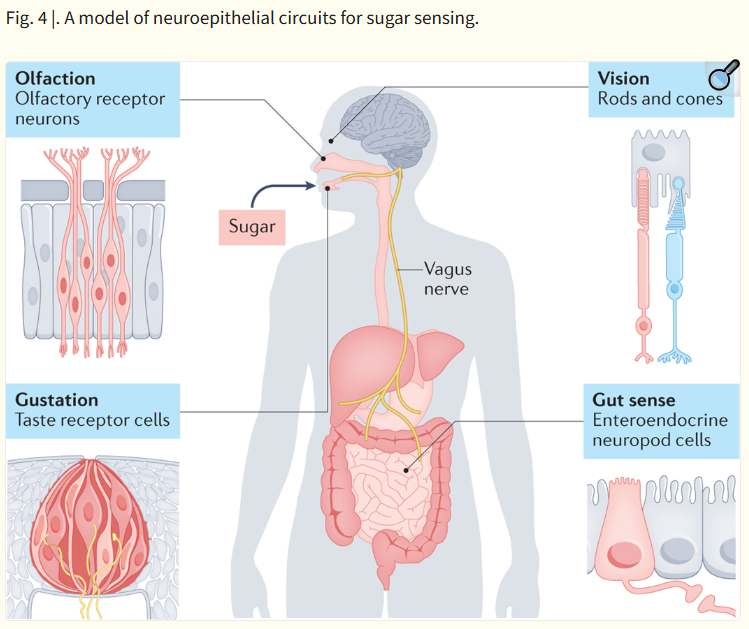
이 그림은
설탕(sugar)을 감지하는 장-뇌, 미각, 후각, 시각의 신경상피 회로(neuroepithelial circuits)를
비교·통합하여 보여주는 모델입니다.
주요 비교
감각 기관 수용체 세포 전달 경로 특징
| Olfaction (후각) | Olfactory receptor neurons | 후각 신경 → 뇌 | 공기 중 냄새 분자 감지 |
| Gustation (미각) | Taste receptor cells | 미각 신경 (VII, IX, X) → 뇌 | 혀에서 단맛 감지 |
| Vision (시각) | Rods and cones | 시신경 → 뇌 | 시각적 정보 |
| Gut sense (장 감각) | Enteroendocrine neuropod cells | Vagus nerve (미주신경) → 뇌 | 장 내 영양소(설탕) 직접 감지 |
핵심 메시지
- 설탕 감지는 혀(미각)뿐만 아니라 장(Gut)에서도 neuropod cells를 통해 미주신경을 거쳐 뇌로 직접 전달됩니다.
- 이는 pre-ingestive(섭취 전, 혀)와 post-ingestive(섭취 후, 장) 감각이 연속적(continuity)이라는 개념을 시각적으로 보여줍니다.
- Gut sense는 neuropod cells가 글루타메이트를 이용해 빠른 신경 신호를 보내는 직접 회로임을 강조.
연구적 의의
- 장-뇌 축(gut-brain axis)이 단순 호르몬 전달이 아닌, 신경상피 회로를 통해 시각·미각·후각과 동등한 수준의 정교한 감각 시스템임을 증명.
- 설탕 선호(sugar preference), 식욕 조절, 중독 기전 연구의 새로운 프레임워크 제공.
https://pmc.ncbi.nlm.nih.gov/articles/PMC9886228/
연구 내용
뇌 보상 회로를 자극해 설탕 선호를 만드는 신경학적 메커니즘을 정리한 2022년 Nature 리뷰 |

| Box 1. Sugar 요약 설탕(Sugar)의 기원과 역사
|
| 이 그림은 장 상피세포(gut epithelial cells)가 포도당(glucose)을 감지하고 신호를 전달하는 3가지 주요 경로를 보여줍니다. ① SGLT1 경로 (Sodium-Glucose Linked Transporter 1)
|

연구 내용
 상단: 전체 개요
1. SGLT1 (소장 상피세포)
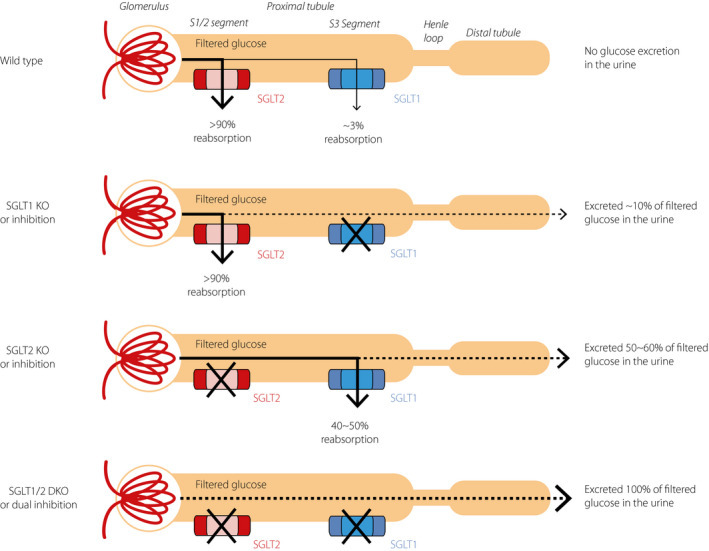
|

연구 핵심
대식세포 M2 극성을 유도해 유방암의 타목시펜 내성을 일으킨다는 2021년 연구 |
연구 내용
 의의
장과 뇌에서 영양을 감지하고 대사·행동 반응을 유발하는 메커니즘을 정리한 2017년 리뷰 |
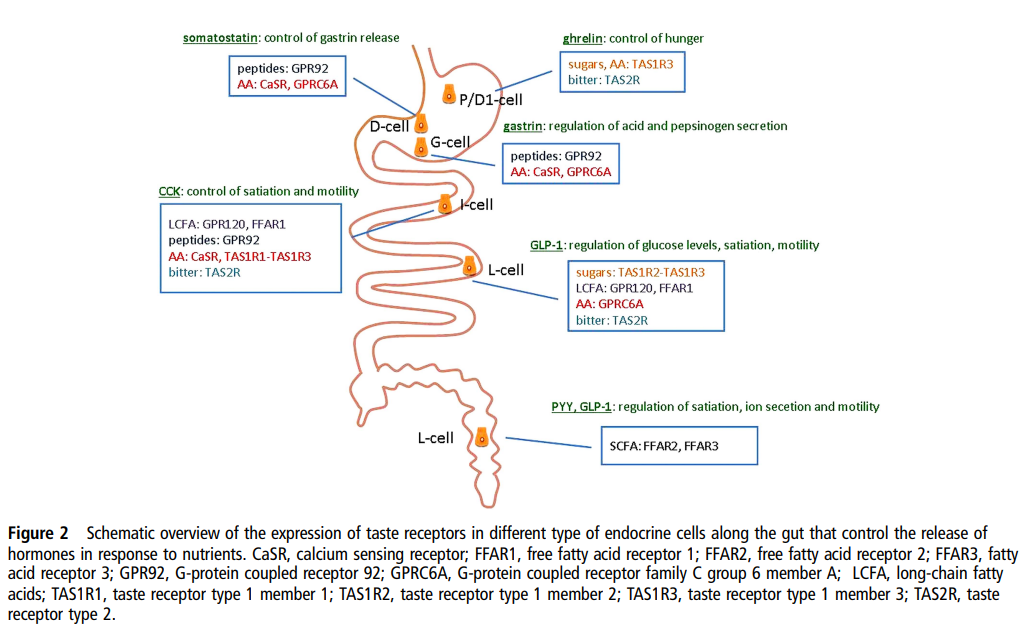
| 이 그림은 장(gut)의 다양한 내분비세포(endocrine cells)에 미각 수용체(taste receptors)가 어떻게 분포하고, 영양소를 감지해 호르몬 분비를 조절하는지 보여줍니다. 주요 세포와 역할
|
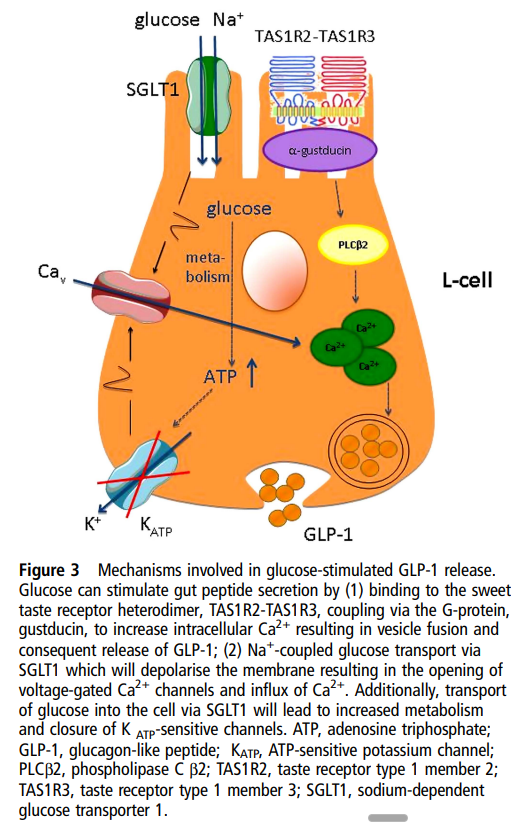
| 이 그림은 L-cell(장 L-세포)에서 포도당(glucose)이 자극하여 GLP-1(Glucagon-like peptide-1)이 분비되는 주요 기전을 보여줍니다. 두 가지 주요 경로 1. Sweet Taste Receptor 경로 (TAS1R2-TAS1R3)
|
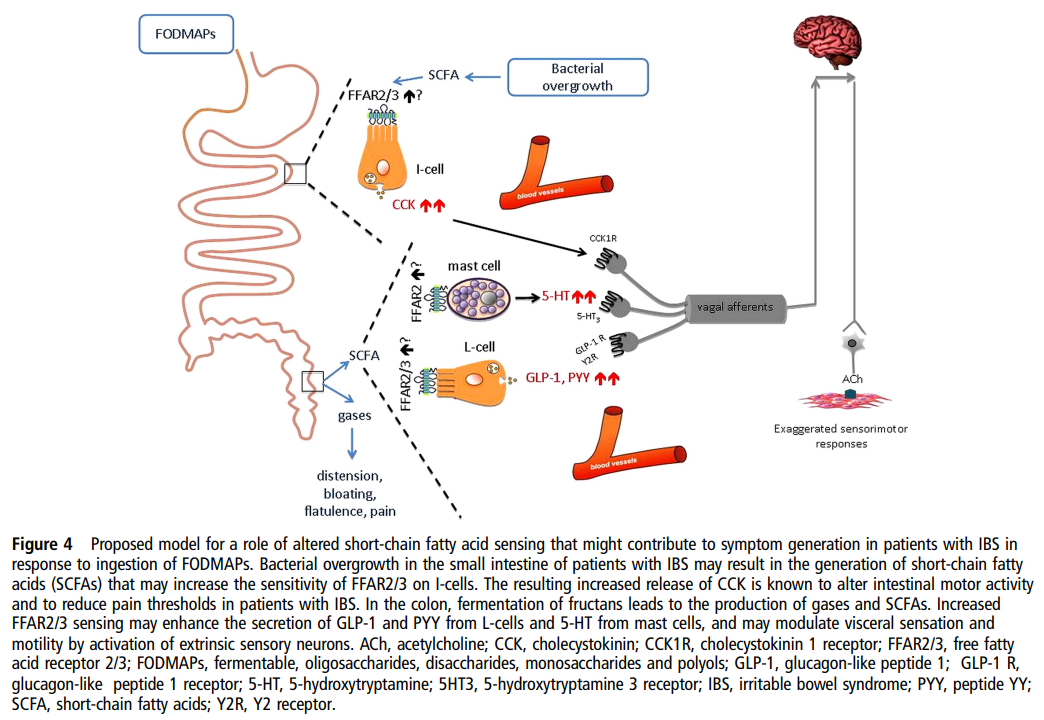
| 주요 기전 흐름 1. 소장에서 Bacterial overgrowth (세균 과증식)
|
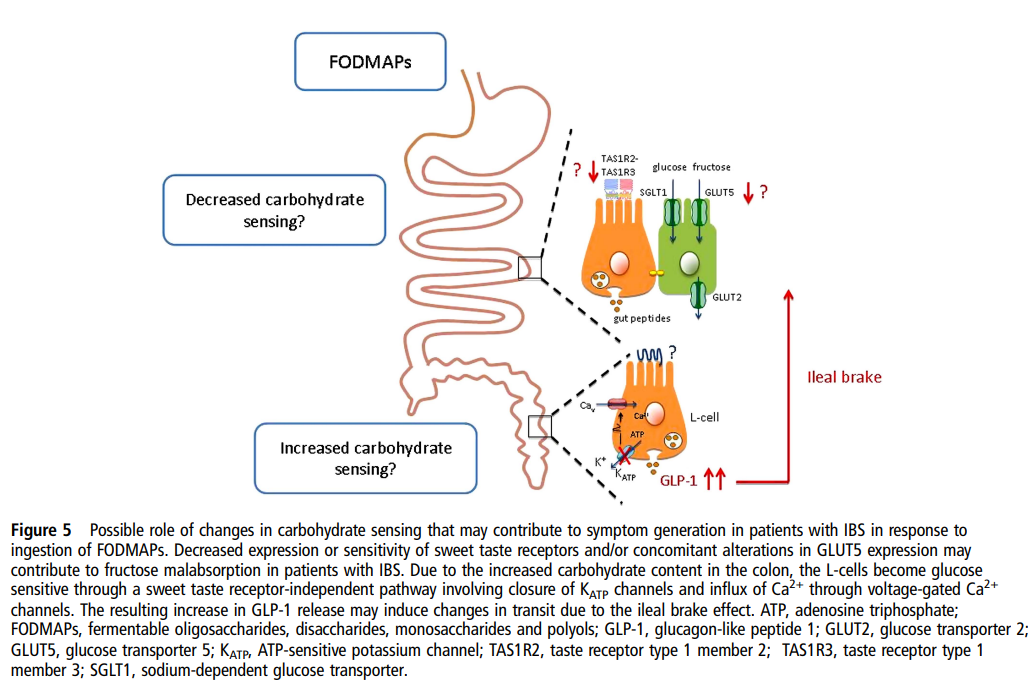
| IBS 환자에서는 단맛 수용체 감수성 저하와 L-세포 과민 반응이라는 이중 이상이 존재하여 FODMAPs에 대한 과민 반응이 증폭된다는 모델 주요 기전 1. Decreased carbohydrate sensing (탄수화물 감지 감소)
|
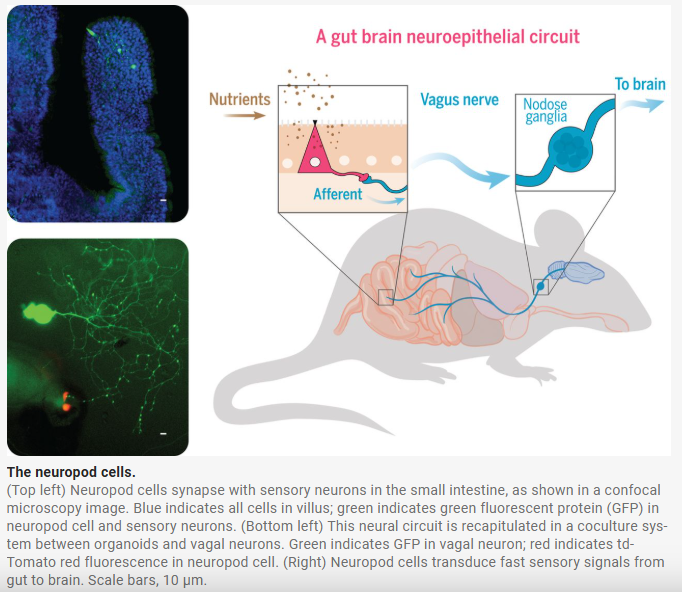
| 이 그림은 2018년 Science 논문(Kaelberer et al.)에서 제시된 장-뇌 직접 연결 회로를 시각적으로 보여주는 핵심 Figure입니다. 주요 발견: Neuropod cells (신경돌기 세포)
장의 neuropod cells가 영양소를 감지하면 글루타메이트를 통해 미주신경을 직접 자극하여 뇌로 빠르게 신호를 보내는 gut-brain neuroepithelial circuit을 발견한 획기적인 연구 |
https://www.science.org/doi/10.1126/science.aat5236
연구의 핵심 발견
장-뇌 축(gut-brain axis)에서
영양소 감지(nutrient sensing)가 어떻게 뇌로 직접 전달되는지 밝힌 획기적인 연구입니다.
기존에는
장에서 분비되는 호르몬(GLP-1 등)이 혈액을 통해 간접적으로 뇌에 신호를 보낸다고 알려져 있었으나,
이 논문은 장 신경세포(glutamatergic enteroendocrine cells)가
글루타메이트(glutamate)를 신경전달물질로 사용하여
미주신경(vagus nerve)을 통해
뇌로 직접 신호를 전달하는 새로운 신경 회로를 최초로 발견했습니다.
주요 내용
- 장 내 영양소 감지 세포: 특정 enteroendocrine cells가 포도당, 아미노산 등의 영양소를 감지.
- 직접 신경 전달: 이 세포들이 글루타메이트를 방출하여 미주신경 구심성 섬유를 자극.
- 뇌로의 신호: 미주신경을 통해 뇌간(nucleus tractus solitarius)으로 빠르게 전달 → 식욕 조절, 포만감, 대사 조절 등.
- 기존 호르몬 경로와 비교: 호르몬 경로는 느리고 간접적인 반면, 이 신경 회로는 매우 빠르고 직접적임.
연구의 의의
- 장-뇌 직접 연결의 존재를 증명 → “gut feelings”의 생물학적 근거 제공.
- 비만, 당뇨, 식이장애 치료에서 미주신경 자극이나 장 신경세포 표적 치료의 새로운 가능성 제시.
- 이후 gut-brain axis 연구의 중요한 전환점이 된 landmark paper.
한 줄 요약:
장에서 영양소를 감지한 세포가
글루타메이트를 이용해 미주신경을 통해
뇌로 직접 신호를 보내는 새로운 gut-brain neural circuit을 발견한 2018년 Science 논문
연구 핵심
설탕 vs 인공감미료 선호 행동을 결정한다는 2022년 Nature Neuroscience 연구 |
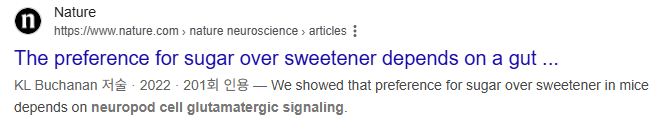
| Box 2 . Neurotransmission from gut to brain. In 2011, an anatomical peculiarity was observed in a subset of enteroendocrine cells that opened a new mechanism for intestinal communication. Some of these sensory cells had a cytoplasmic protrusion at the base of the cells which resembled a neuronal axon171. Subsequent work confirmed that some enteroendocrine cells, both with and without a basal extension, could from synapses with enteric, vagal and splanchnic nerves66,97,172,173. To account for the neuron-like function in these cells, the term neuropod cell was extended to those epithelial cells engaging in neurotransmission66. This gut neuroepithelial sensory circuit could serve to account for the chemical, mechanical and thermal properties of ingested nutrients. The similarity between neuroepithelial circuits in the gastrointestinal tract and other sensory systems like those of taste, olfaction and vision raised the hypothesis that there exists continuity between pre-ingestive and post-ingestive sensory mechanisms. Neuropod cells are a subset of enteroendocrine cells that are differentiated by their neural function. Transcript and protein analyses have shown that subpopulations of enteroendocrine cells express pre-synaptic proteins such as synapsin 1, piccolo, bassoon, MUNC13B, RIMS2, latrophilin 1 and neurexin 297. Ultrastructural investigations have revealed that neuropods contain neurofilaments (see the figure, part a) as well as both dense core vesicles containing peptides and synaptic clear vesicles containing neurotransmitters (see the figure, part b)164. In vitro, neuropod cells synapse with co-cultured neurons, and optogenetic activation of neuropod cells triggers excitatory postsynaptic potentials in connected neurons within 60 milliseconds66. In vivo, monosynaptic tracing using rabies virus demonstrated that small intestinal neuropod cells form monosynaptic connections with vagal neurons to rapidly communicate nutrient sensory information to the brain (see the figure, part c)66. In a series of studies utilizing organoid cultures, Bellono et al. showed that colonic enterochromaffin cells synapse with peripheral nerves to rapidly transduce the presence of irritants using serotonin172. Subsequent results showed that these cells could also form purinergic synapses173. In vivo, luminal optogenetic manipulation of neuropod cells shows that these cells are required for sugar preference99. Parts a and b adapted with permission from REF.164. Part c adapted with permission from REF.66. Neuropod cells는 enteroendocrine cells(장내분비세포)의 특수 아형으로, 신경세포와 유사한 기능을 가진 세포입니다. 2011년 처음 발견된 기저부 cytoplasmic protrusion(신경돌기 같은 구조)을 가진 세포가 그 시작입니다. 주요 특징
|
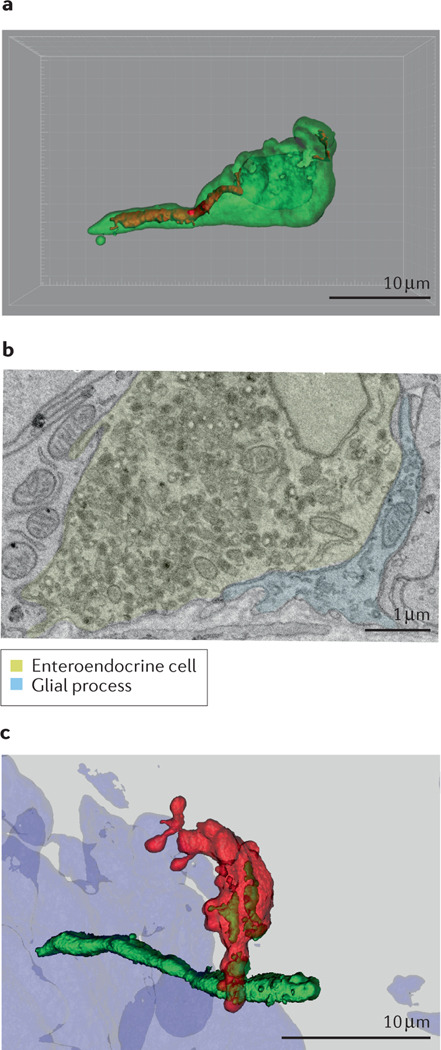
a. Vagal neurons의 Subpopulations (아형)
영양소·기계적·삼투압을 각각 또는 조합으로 감지하는 다양한 subpopulation으로 구성되어 있음. b. Vagal nodose responses to gastrointestinal stimuli
|
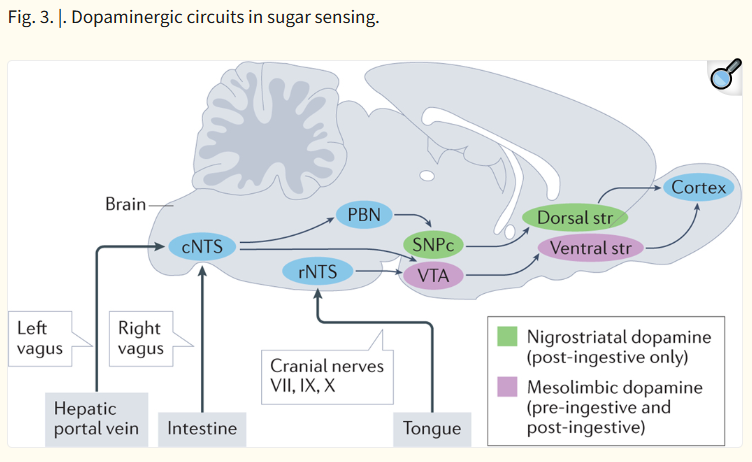
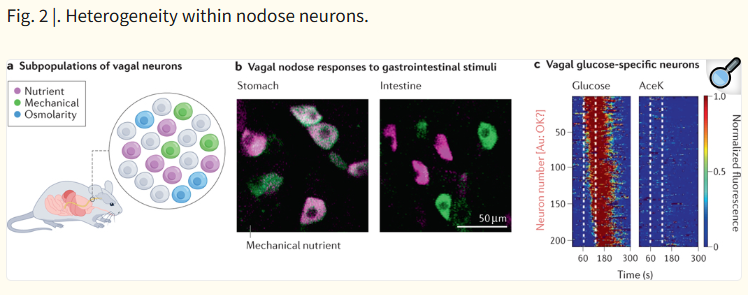
| 이 그림은 설탕(당) 감지 과정에서 도파민(dopamine)이 관여하는 뇌 회로를 보여줍니다. 특히 pre-ingestive(섭취 전)와 post-ingestive(섭취 후) 단계에서 도파민 경로가 다르게 작용한다는 점을 강조합니다. 주요 회로 구성 1. 입력 경로 (Input)
설탕은 혀·장·간문맥에서 감지되어 vagus nerve → NTS → VTA/SNpc를 통해 mesolimbic (pre/post)과 nigrostriatal (post) 도파민 회로를 활성화시켜 강력한 보상 반응을 유발한다는 모델 |
Nat Rev Neurosci
. Author manuscript; available in PMC: 2023 Apr 1.
Published in final edited form as: Nat Rev Neurosci. 2022 Jul 25;23(10):584–595. doi: 10.1038/s41583-022-00613-5
The neural basis of sugar preference
Winston W Liu 1,2,3, Diego V Bohórquez 1,2,3,*
- Author information
- Article notes
- Copyright and License information
PMCID: PMC9886228 NIHMSID: NIHMS1827107 PMID: 35879409
The publisher's version of this article is available at Nat Rev Neurosci
Abstract
When it comes to food, one tempting substance is sugar. Although sweetness is detected by the tongue, the desire to consume sugar arises from the gut. Even when sweet taste is impaired, animals can distinguish sugars from non-nutritive sweeteners guided by sensory cues arising from the gut epithelium. Here, we review the molecular receptors, cells, circuits and behavioural consequences associated with sugar sensing in the gut. Recent work demonstrates that some duodenal cells, termed neuropod cells, can detect glucose using sodium glucose co-transporter 1, and release glutamate onto vagal afferent neurons. Based on these and other data, we propose a model in which specific populations of vagal neurons relay these sensory cues to distinct sets of neurons in the brain, including neurons in the caudal nucleus of the solitary tract, dopaminergic reward circuits in the basal ganglia, and homeostatic feeding circuits in the hypothalamus that alter current and future consumption. This emerging model highlights the critical role of the gut in sensing the chemical properties of ingested nutrients to guide appetitive decisions.
음식과 관련하여
가장 유혹적인 물질 중 하나는 설탕입니다.
단맛은 혀로 감지되지만,
설탕을 섭취하고자 하는 욕구는 장(gut)에서 비롯됩니다.
단맛 감각이 손상된 경우에도 동물들은
장 상피에서 발생하는 감각 신호를 통해 설탕과 비영양성 감미료를 구분할 수 있습니다.
본 리뷰에서는
장(gut)에서의 설탕 감지와 관련된 분자 수용체, 세포, 회로, 그리고 행동적 결과를 종합적으로 검토합니다.
최근 연구에 따르면,
십이지장(duodenum)의 특정 세포인 neuropod cells는
SGLT1(나트륨-포도당 공동수송체 1)을 이용해 포도당을 감지하고,
미주신경 구심성 신경(vagal afferent neurons)에 글루타메이트를 방출합니다.
이러한 연구와 기타 데이터를 바탕으로,
우리는 특정 vagal neuron 집단이
이러한 감각 신호를 뇌의 서로 다른 뉴런 집단으로 전달하는 모델을 제안합니다.
여기에는
caudal nucleus of the solitary tract (cNTS)의 뉴런,
기저핵(basal ganglia)의 도파민 보상 회로,
그리고 시상하부(hypothalamus)의 항상성 섭식 회로가 포함되며,
이는 현재와 미래의 식이 행동을 변화시킵니다.
이 새롭게 부상하는 모델은
섭취된 영양소의 화학적 특성을 감지하여
식욕(apetitive) 결정을 안내하는 데
장(gut)이 핵심적인 역할을 한다는 점을 강조합니다
Introduction
We seek nourishment guided by our senses, and when it comes to food, they determine which are nutritious, safe, satisfying, palatable and pleasurable. Although pursuing a particular diet is a conscious process, daily food intake is largely a product of subconscious preferences and food availability. Following the agricultural revolution, food availability increased exponentially, freeing a system once used mainly for survival to also allow for the pursuit of pleasure1,2.
Pleasurable foods, like sugar extracted from sugar cane or beets, became common crops that could be processed, preserved, and promoted in the food chain. Sugar, which was once consumed as part of natural foods such as fruit, could now be distilled into its pure form. Industrial food processing further dissociated it from sensory cues. It was now possible to produce sweeteners that lack nutritive value. But our bodies could not adapt rapidly enough to interpret such artificial stimuli leading to skewed nutritive choices followed by a steep rise in obesity and metabolic disease3. Efforts to target the taste of foods alone through non-nutritive artificial sweeteners[G] have failed to curb disorders of food intake, revealing a pressing need to understand how animals detect, distinguish and choose to consume sugar4,5.
Our diet is broadly monitored by two categories of senses: the pre-absoprtive and the post-absorptive. The pre-absorptive senses, often referred to as exteroceptive[G], monitor food before it crosses an epithelial surface of the body. In this category are the cephalic senses of olfaction and gustation as well as the more hidden sense of the gastrointestinal epithelium6. The post-absorptive senses, or interoceptive[G], monitor sensory cues from absorbed nutrients reaching the liver, pancreas, and other long-term energy reporters in adipose and muscular tissues. Both pre-absorptive and post-absorptive senses include conscious and subconscious components. It is the synergy between pre-absorptive and post-absorptive sensory cues that drives behaviour by allowing the brain to create a representation of foods from their appearance to their metabolic value.
This Review details the neural basis of our preference for sugar (Box 1) — from its pre-absorptive detection to its ability to drive behavioural preferences. The functions of gustation in recognizing sugar are well reviewed7–9. Instead, this Review focuses on the role of the gut in recognizing glucose to drive behavioural preference. Novel tools first developed for use in the CNS are now being adapted for use in the periphery, which has accelerated our understanding of gut–brain signaling. Emerging evidence suggests that the brain receives rapid sensory cues from the gut independently of the oral cavity, and that these subconscious signals may subvert conscious processes in the preference for sugars.
우리는
감각에 이끌려 영양분을 추구하며,
음식의 경우 감각은 어떤 것이 영양가 있고, 안전하며, 만족스럽고, 맛있고, 즐거운지를 결정합니다.
특정 식이를 추구하는 것은 의식적인 과정이지만,
일상적인 음식 섭취는 대부분 무의식적인 선호와 음식 가용성의 결과물입니다.
농업 혁명 이후 음식 가용성이 기하급수적으로 증가하면서,
한때 생존을 위한 시스템이었던 것이 쾌락 추구까지 가능하게 되었습니다¹,².
설탕수수나 사탕무에서 추출한 설탕과 같은 즐거운 음식은
가공·보존·유통이 가능한 일반 작물이 되었고,
한때 과일 같은 자연 식품의 일부로 소비되던 설탕은 이제 순수한 형태로 정제될 수 있게 되었습니다.
산업적 식품 가공은
설탕을 감각 신호로부터 더욱 분리시켰습니다.
이제
영양가가 없는 감미료를 생산하는 것도
가능해졌습니다.
그러나
우리 몸은 이러한 인공적 자극을 해석할 만큼 빠르게 적응하지 못했고,
이는 영양 선택의 왜곡을 초래하여 비만과 대사 질환의 급격한 증가를 가져왔습니다³.
비영양성 인공 감미료만을 이용해 음식의 맛만을 표적화하려는 노력은
식이 장애를 줄이는 데 실패했으며,
동물들이 설탕을 어떻게 감지하고, 구분하며, 선택적으로 섭취하는지를 이해해야 할 절실한 필요성을 드러냈습니다⁴,⁵.
우리의 식이는
크게 두 가지 범주의 감각에 의해 감시됩니다:
흡수 전(pre-absorptive)과
흡수 후(post-absorptive) 감각입니다.
흡수 전 감각(종종 외수용성(exteroceptive)이라고 함)은
음식이 신체의 상피 표면을 통과하기 전에 감시합니다.
이 범주에는
후각(olfaction)과 미각(gustation) 같은 두부 감각(cephalic senses)과,
위장관 상피의 보다 숨겨진 감각이 포함됩니다⁶.
흡수 후 감각(내수용성(interoceptive))은
흡수된 영양소가 간, 췌장, 지방 조직, 근육 조직 등 장기 에너지 감지 기관에 도달한 후의 감각 신호를 감시합니다.
흡수 전과 흡수 후 감각 모두
의식적 및 무의식적 요소를 포함합니다.
흡수 전과 흡수 후 감각 신호의 시너지가
음식의 외관부터 대사적 가치까지
뇌가 음식을 대표(representation)하게 만들어 행동을 유도합니다.
본 리뷰는
설탕에 대한 우리의 선호의 신경학적 기초(Box 1 참조) — 흡수 전 감지부터 행동적 선호 유도까지 — 를
자세히 다룹니다.
설탕을 인식하는 미각(gustation)의 기능은
이미 잘 검토되었습니다⁷⁻⁹.
대신,
본 리뷰는 장(gut)이 포도당을 인식하여 행동적 선호를 유도하는 역할에 초점을 맞춥니다.
중추신경계(CNS)에서 처음 개발된 새로운 도구들이
이제 말초(장)로 적용되면서
장-뇌 신호 전달(gut–brain signaling)에 대한 이해가 가속화되었습니다.
최근 증거에 따르면,
뇌는 구강(oral cavity)과 독립적으로 장으로부터 빠른 감각 신호를 받고 있으며,
이러한 무의식적 신호가 설탕 선호에서 의식적 과정을 잠재적으로 무시(subvert)할 수 있습니다.
Box 1 |. Sugar.
The English word ‘sugar’ traces its origin to the Indian subcontinent. The sanskrit word ‘sharkara’, which characterized the ‘gravel-like’ texture of sugar, travelled with Alexander the Great through the Arabic word “sukkar”, and finally landed in Western European words like “sucre” (France) and “azucar” (Spain)165. The earliest recorded use of sugar products arises even earlier, at the time of the agricultural revolution in ~8000 BCE. In fact, sugarcane is purported to be one of the first domesticated crops, and Papua New Guinean nomads were known to chew on raw sugarcane166. The appeal of sugar has evolutionary roots in the fruit-based diet of great apes. The natural sugar in fruit likely served as an anticipatory cue for caloric density167,168.
Sugar generally refers to the chemical compound sucrose, which is produced by plants through photosynthesis169. Sucrose is a disaccharide formed by one glucose and one fructose molecule linked via an ether bond. Cleavage of the bond via the enzyme sucrase-isomaltase is required for absorption. Expression of the enzyme has been detected in the oral cavity in saliva, on taste receptor cells, and more broadly along the brush border of the duodenum and jejunum of the small intestine. Once cleaved, glucose and fructose are absorbed via different transporters on the apical membrane of the intestine. Glucose is subsequently transported into the bloodstream, where pancreatic hormones tightly regulate its concentration. Importantly, glucose is the primary energy source for the CNS and is stored as glycogen in much lower quantities than fats. Thus, the body must continuously replenish glucose stores through gluconeogenesis or dietary intake170. Fructose is then transported to the liver via the bloodstream and undergoes fructolysis before entering gluconeogenesis or lipogenesis.
영어 단어 ‘sugar’는
인도 아대륙에서 유래했습니다.
설탕의 ‘자갈 같은(gravel-like)’ 질감을 나타내던 산스크리트어 ‘sharkara’가
알렉산더 대왕을 따라 아랍어 “sukkar”를 거쳐,
서유럽 언어인 프랑스어 “sucre”와 스페인어 “azucar”로 전파되었습니다¹⁶⁵.
설탕 제품의 가장 오래된 기록은
농업 혁명 시기인 기원전 약 8000년경까지 거슬러 올라갑니다.
실제로 사탕수수는
최초로 재배된 작물 중 하나로 알려져 있으며,
파푸아뉴기니 유목민들은 생 사탕수수를 씹어 먹는 것으로 유명했습니다¹⁶⁶.
설탕의 매력은
대형 유인원의 과일 중심 식이에 진화적 뿌리를 두고 있습니다.
과일에 들어 있는 천연 설탕은
칼로리 밀도(caloric density)에 대한 예고 신호(anticipatory cue)로 작용했을 가능성이 높습니다¹⁶⁷,¹⁶⁸.
설탕(sugar)은 일반적으로
식물이 광합성을 통해 생산하는 화학 물질 수크로스(sucrose)를 가리킵니다¹⁶⁹.
수크로스는
포도당 1분자와 과당 1분자가 에테르 결합(ether bond)으로 연결된 이당류(disaccharide)입니다.
이 결합은
sucrase-isomaltase 효소에 의해 절단되어야 흡수가 가능합니다.
이 효소는
구강(타액), 미각 수용체 세포, 그리고
십이지장과 공장의 brush border를 따라 광범위하게 발현되는 것으로 확인되었습니다.
절단된 후 포도당과 과당은
장 상피세포의 apical membrane에 있는 서로 다른 수송체를 통해 흡수됩니다.
포도당은
혈류로 들어간 후 췌장 호르몬에 의해 그 농도가 정밀하게 조절됩니다.
특히 포
도당은 중추신경계(CNS)의 주요 에너지원이며,
지방에 비해 글리코겐 형태로 저장되는 양이 매우 적습니다.
따라서
몸은 gluconeogenesis(포도당 신생합성)이나
식이를 통해 지속적으로 포도당을 보충해야 합니다¹⁷⁰.
과당은
혈류를 통해 간으로 이동한 후 fructolysis를 거쳐
gluconeogenesis나 lipogenesis(지방 생성) 경로로 들어갑니다.
Behavioural regulation of sugar intake
In 1947, Edward Adolph fed rats diets diluted with indigestible agents such as cellulose. He found that while the total amount of food consumed by the rats increased with dilute diets, the total amount of calories consumed remained the same10. This observation first associated the quantity of food consumed to its nutritional content; in other words, animals eat to meet caloric requirements. However, in the 1950s, Curt Richter observed that rats selectively increased intake of carbohydrates when challenged with the removal of the adrenal glands or the pancreas11. These studies demonstrated that animals can also adjust their desire to seek out specific foods in response to the properties of ingested nutrients.
In 1952, rats with stomach fistulas were used to postulate that a solution “injected directly into the stomach serves as a reward to produce learning”12. This role of gastrointestinal signalling in the behavioural regulation of food intake was substantiated in 1969 when Garvin Holman published a classic study in which they gave rats flavoured saccharin solutions paired with intragastric infusions of nutrient-rich eggnog or water13. When the rats were later given a choice between the two flavoured solutions in the absence of an intragastric infusion, they preferred the solution originally paired with nutrients. In the ensuing decades, other studies refined the techniques to further substantiate the hypothesis that the brain receives rapid signals from food stimuli detected by specific receptors in the gastrointestinal tract14.
A primary level of behavioural regulation of food intake requires animals to discriminate between different nutrients, assign value to nutrients depending on their utility, and decide whether or not to continue their consumption. Thus, nutrients entering the gut should be able to both positively and negatively reinforce consumption. The ability of nutritive substances to reduce consumption has been well documented through studies on satiation[G] and satiety[G]15–17. However, more recent work has focused on positive reinforcement[G], which has been termed ‘appetition[G]’18. These processes are difficult to probe even in animal models, and most experimental paradigms require a training phase in which mice learn to respond for the infusion of intraluminal stimuli. For sugars, trained mice increase their licking responses towards an empty spout, which triggers intragastric self-infusion of glucose over water19,20. Thus, mice will perform operant[G] tasks such as licking a dry spout to self-administer intra-intestinal infusions of glucose21,22. Notably, not all carbohydrates can sustain operant behaviour to the same extent: the monosaccharide fructose is much weaker at doing so than glucose or sucrose19.
A secondary level of food intake regulation requires learning nutrient value for future decision-making. The animal must develop a preference, recall that preference, and act on it. This function of positive reinforcement of foods in associative learning has been extensively characterized using flavour–nutrient conditioning[G] paradigms, in which mice are trained to associate flavoured solutions (conditioned stimulus) with intragastric infusions of various nutrients (unconditioned stimulus). Then, in a separate session, mice select between the flavoured solutions. Using this experimental approach, it was found that the set of carbohydrates that can condition flavour preferences mirror those that can induce operant behaviour. For instance, glucose as well maltose and sucrose can strongly condition a preference. Maltose is made of two glucose molecules whereas sucrose is a mix of glucose and fructose. Contrary to glucose, intragastric infusion of fructose is less or completely ineffective at conditioning a flavour preference23–26. The conditioned preference learning is both rapid and persistent. With strong conditioned stimuli, rats can acquire robust conditioned flavour preferences after a single conditioning day. In one study adopting such an approach, after training, a single 6 ml intragastric glucose infusion produced a long-lasting preference memory, lasting for at least 13 days post-conditioning27.
The timescales of these primary and secondary levels of food intake regulation appear to be different: whereas acceptance of a solution occurs within seconds to minutes of a meal, learning probably takes place over hours across meals. Although conditioned learning has not been assessed for intervals shorter than 24 hours, the gastrointestinal tract can distinguish between specific carbohydrates within secondsREF99. Over the last 75 years since Adolph began feeding cellulose to rats, it has become clear that the gastrointestinal tract can distinguish between nutrient stimuli and guide appetitive behaviors.
Behavioural regulation of sugar intake (설탕 섭취의 행동 조절)
1947년 Edward Adolph는
쥐에게 섬유소(cellulose)와 같은 소화 불가능한 물질로 희석된 사료를 먹였습니다.
그는
희석된 사료에서는 쥐가 섭취하는 총 음식량은 증가했지만,
총 칼로리 섭취량은 일정하게 유지된다는 것을 발견했습니다¹⁰.
이 관찰은
처음으로 섭취하는 음식의 양이 그 영양 성분(칼로리)과 연관되어 있다는 점을 밝혔습니다.
즉, 동물은
칼로리 요구량을 충족시키기 위해 먹는다는 것입니다.

| 주요 내용 및 실험 설계 Richter는 부신절제술(adrenalectomy)을 한 쥐를 이용해 실험했습니다.
|
그러나
1950년대 Curt Richter는
부신(adrenal glands)이나 췌장(pancreas)을 제거한 쥐가
탄수화물 섭취를 선택적으로 증가시킨다는 것을 관찰했습니다¹¹.
이러한 연구들은
동물이 섭취된 영양소의 특성에 따라
특정 음식을 찾는 욕구를 조절할 수 있음을 보여주었습니다.
1952년에는
위 누공(stomach fistula)이 있는 쥐를 이용해
“위에 직접 주입된 용액이 학습을 유도하는 보상(reward)으로 작용한다”는 가설을 제기했습니다¹².
위장관 신호가 음식 섭취의 행동 조절에 미치는 역할은
1969년 Garvin Holman의 고전적 연구로 확고히 입증되었습니다.
이 연구에서
쥐에게 향이 첨가된 saccharin 용액을 영양이 풍부한 eggnog 또는 물의 위내 주입(intragastric infusion)과 짝지어 주었습니다¹³.
이후
쥐가 위내 주입 없이 두 향이 있는 용액을 선택하게 했을 때,
영양소와 짝지어졌던 용액을 선호했습니다.
그 후 수십 년 동안 다른 연구들은
위장관 내 특정 수용체가 감지한 음식 자극으로부터 뇌가 빠른 신호를 받는다는 가설을
더욱 정교하게 검증하는 기술을 발전시켰습니다¹⁴.
음식 섭취 행동 조절의 1차 수준은
동물이 서로 다른 영양소를 구별하고,
그 영양소의 효용(utility)에 따라 가치를 부여하며,
계속 섭취할지 여부를 결정하는 것입니다.
따라서
장으로 들어가는 영양소는
긍정적 강화(positive reinforcement)와 부정적 강화(negative reinforcement) 모두를 할 수 있어야 합니다.
영양 물질이 섭취를 감소시키는 능력은
satiation(만족)과 satiety(포만)에 대한 연구를 통해 잘 알려져 있습니다¹⁵⁻¹⁷.
그러나
최근 연구는 긍정적 강화(positive reinforcement),
즉 ‘appetition’¹⁸이라고 불리는 현상에 초점을 맞추고 있습니다.
이러한 과정은
동물 모델에서도 탐색하기 어렵고,
대부분의 실험 패러다임은 쥐가 장 내 자극(intraluminal stimuli) 주입을 위해 반응하는 훈련 단계를 요구합니다.
설탕의 경우,
훈련된 쥐는 빈 spout(주둥이)을 핥는 행동을 증가시켜 포도당의 자가 주입을 유발합니다¹⁹,²⁰.
따라서
쥐는 dry spout(빈 주둥이)을 핥는 operant task(조작적 과제)를 수행하여
장 내 포도당 주입을 스스로 투여합니다²¹,²².
주목할 점은
모든 탄수화물이 동일한 정도로 operant 행동을 유지할 수 있는 것은 아니라는 것입니다:
단당류인 fructose(과당)는 glucose나 sucrose에 비해 훨씬 약합니다¹⁹.
음식 섭취 조절의 2차 수준은
미래 의사결정을 위한 영양소 가치 학습을 요구합니다.
동물은
선호(preference)를 형성하고,
그 선호를 기억하며, 그에 따라 행동해야 합니다.
이러한 flavour–nutrient conditioning(향-영양소 조건화) 패러다임은
긍정적 강화의 기능을 광범위하게 규명하는 데 사용되었습니다.
쥐에게 향이 첨가된 용액(조건 자극)과 다양한 영양소의 위내 주입(무조건 자극)을 짝지어 훈련시킨 후,
별도의 세션에서 향이 있는 용액들 사이에서 선택하게 합니다.
이 실험 접근법을 통해
flavour preference를 조건화할 수 있는 탄수화물 집단이
operant 행동을 유도할 수 있는 집단과 일치한다는 것이 밝혀졌습니다.
예를 들어,
glucose, maltose, sucrose는
강력한 preference를 조건화할 수 있습니다.
Maltose는
포도당 2분자로, sucrose는 포도당과 과당의 혼합물입니다.
Glucose와 달리
fructose의 위내 주입은 flavour preference 조건화에 덜 효과적이거나 완전히 무효입니다²³⁻²⁶.
이러한 conditioned preference 학습은
빠르고 지속적입니다.
강한 조건 자극에서는
쥐가 단 하루의 conditioning만으로도 강력한 conditioned flavour preference를 획득할 수 있습니다.
한 연구에서는
conditioning 후 단 6ml의 위내 glucose 주입만으로도 최소 13일 동안 지속되는 장기 선호 기억이 형성되었습니다²⁷.
이러한 1차 및 2차 수준의 음식 섭취 조절 시간 scale은 다릅니다:
용액 수용(acceptance)은
식사 후 초~분 단위로 일어나지만,
학습은 수 시간~수 일에 걸쳐 진행됩니다.
conditioned learning은
24시간 미만의 간격에서는 평가되지 않았지만,
위장관은 몇 초 만에 특정 탄수화물을 구분할 수 있습니다⁹⁹.
Adolph가 쥐에게 섬유소를 먹이기 시작한 지 75년이 지난 지금,
위장관이 영양 자극을 구분하고 식욕 행동(appetitive behaviors)을 안내할 수 있다는 것이 분명해졌습니다.
Molecular sugar detectors
Detection of sucrose and its byproducts by epithelial cells along the contiguous sensory surface from tongue to intestine occurs via three primary pathways: taste receptors (comprising taste receptor type 1 member 2 (T1R2) and T1R3 subunits), glucose transporters (for example, sodium/glucose cotransporter 1 (SGLT1)) and ATP-gated potassium (KATP) channels. These three mechanisms are shared across various tissues in both the pre-absorptive and post-absorptive sensory systems. Outside of the oral cavity, taste receptors are also expressed in the pancreas28,29, adipocytes30,31 and the brain32,33. Similarly, SGLTs are not restricted to the intestine, and are also expressed in the tongue34, pancreas35, brain36 and hepatic portal system37. Within the gastrointestinal tract, T1R2–T1R3 receptors and SGLT1 occur along the apical surface facing the lumen of the gastrointestinal tract38,39, whereas KATP channels detect the end products of glucose metabolism within the cell (Fig. 1)40.
Molecular sugar detectors (설탕의 분자 감지기)
수크로스와 그 분해 산물은
혀에서 장까지 연속된 감각 표면의 상피세포에서 세 가지 주요 경로를 통해 감지됩니다:
맛 수용체(T1R2-T1R3),
포도당 수송체(SGLT1),
그리고 ATP-의존성 칼륨(KATP) 채널입니다.
이 세 가지 기전은
흡수 전(pre-absorptive)과 흡수 후(post-absorptive) 감각계의 다양한 조직에서 공유됩니다.
구강 외에도 맛 수용체는
췌장²⁸,²⁹, 지방세포³⁰,³¹, 뇌³²,³³에서도 발현됩니다.
SGLT 역시
장에 국한되지 않고 혀³⁴, 췌장³⁵, 뇌³⁶, 간문맥계³⁷에서도 발현됩니다.
위장관 내에서 T1R2–T1R3 수용체와 SGLT1은
장 내강(lumen)을 향한 apical membrane에 위치하며³⁸,³⁹,
KATP 채널은 세포 내에서 포도당 대사의 최종 산물을 감지합니다(Fig. 1)⁴⁰.
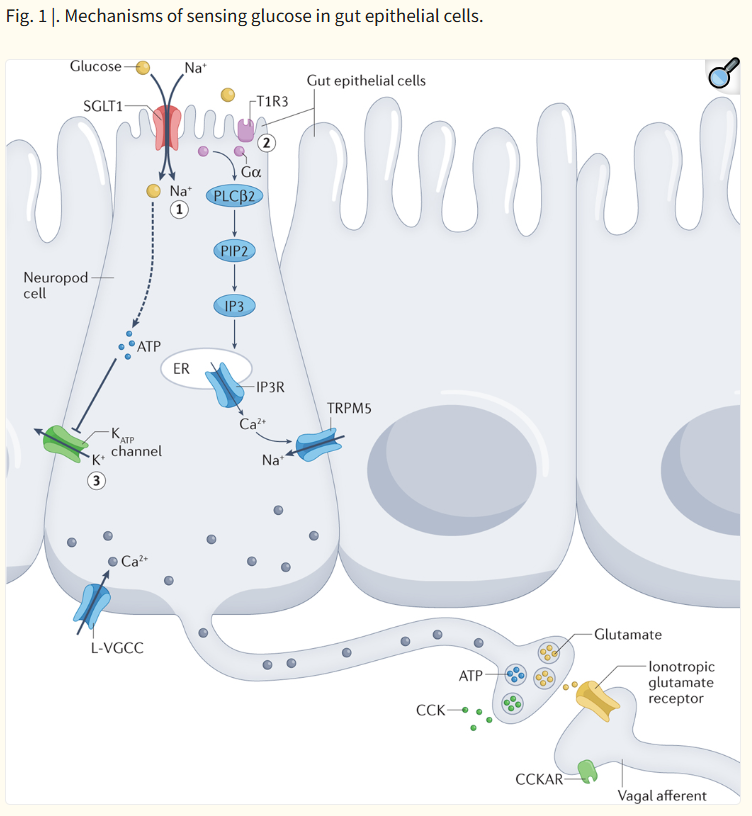
Fig. 1 |. Mechanisms of sensing glucose in gut epithelial cells.
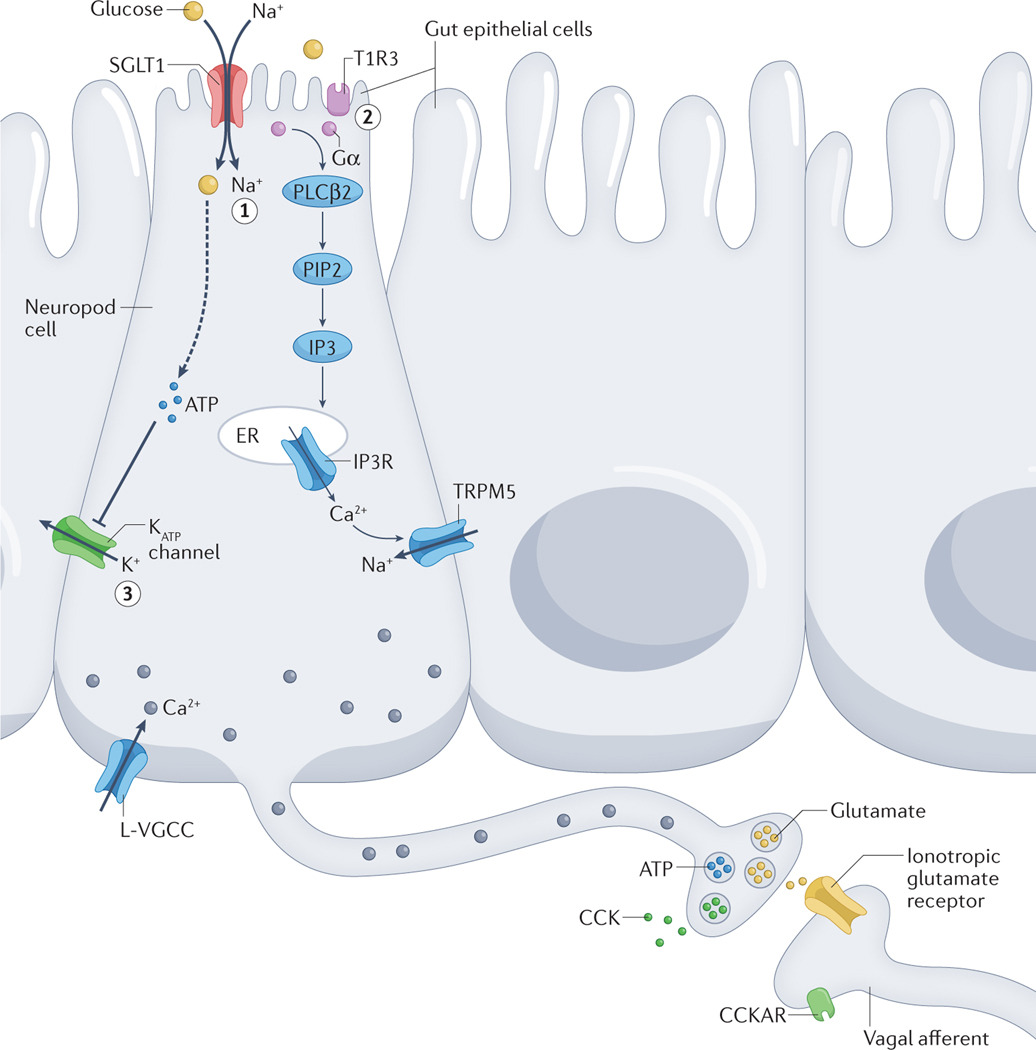
In intestinal epithelial cells known as neuropod cells, glucose can lead to cellular depolarization and subsequent signalling molecule release in three ways. Sodium glucose co-transporter 1 (SGLT1)-dependent inflow of Na+ drives glucose absorption and leads to depolarization. L-type voltage-gated calcium channels (L-VGCCs) are then hypothesized to be activated, which triggers a rise in intracellular Ca2+ levels and the release of glutamate (1). Activation of T1R3-subunit containing taste receptors by glucose triggers a secondary signalling cascade involving the following: α-gustducin (Gα), phospholipase Cβ2 (PLCβ2), phosphatidylinositol 4,5-bisphosphate (PIP2), inositol 1,4,5-trisphosphate (IP3), IP3 receptors (IP3Rs) on the endoplasmic reticulum (ER), and TRPM5 (2). Depolarization is hypothesized to induce voltage-gated CAHLM1–CAHLM3 channels to release ATP non-vesicularly (not shown). VGCCs can also be activated that lead to increased intracellular Ca2+ levels and subsequent vesicular release of ATP. ATP-gated potassium (KATP) channels close in response to the end-products of glucose metabolism increasing the ATP:ADP ratio (3). VGCCs can then open that trigger the release of hormones such as cholecystokinin (CCK). CCKAR, CCK A receptor.
장 상피세포(neuropod cells)에서 포도당은
세포 탈분극(cellular depolarization)과 후속 신호분자 방출을
세 가지 방식으로 유발할 수 있습니다.
- Sodium glucose co-transporter 1 (SGLT1) 의존적 Na⁺ 유입: Na⁺와 함께 포도당이 흡수되면서 탈분극이 발생합니다. 이후 L-type voltage-gated calcium channels (L-VGCCs)가 활성화되어 세포 내 Ca²⁺ 농도가 증가하고, glutamate 방출이 유발됩니다.
- T1R3 subunit을 포함한 taste receptor 활성화: 포도당이 T1R3를 포함한 단맛 수용체를 활성화하면 α-gustducin (Gα), phospholipase Cβ2 (PLCβ2), phosphatidylinositol 4,5-bisphosphate (PIP2), inositol 1,4,5-trisphosphate (IP3), endoplasmic reticulum (ER)상의 IP3 receptors (IP3Rs), TRPM5을 포함하는 2차 신호 전달 경로가 활성화됩니다.
- KATP channel 경로: 포도당 대사 산물에 의해 ATP:ADP 비율이 증가하면 ATP-gated potassium (KATP) channels이 닫히면서 탈분극이 발생합니다. 이로 인해 voltage-gated calcium channels (VGCCs)이 열리고, cholecystokinin (CCK) 같은 호르몬의 방출이 유발됩니다. (CCKAR: CCK A receptor)
Sweet taste receptors
Sweet[G] taste receptor subunits were first identified by three independent groups and have been primarily studied in the context of taste receptor cells of the tongue41–43. As indicated above, sweet taste is conveyed by a heterodimer of class C G-protein coupled receptors (GPCRs), namely T1R2 and T1R343,44. These GPCRs contain four major domains: an extracellular venus-flytrap domain, a cysteine-rich linker domain, a transmembrane seven-helix domain and an intracellular domain that interacts with G proteins. Based on homology to other receptors and modelling, it is predicted that sugar binding to the venus-flytrap domain triggers a conformational shift that closes the two flytrap lobes and activates the intracellular domain to bind its G protein45,46. Artificial sweeteners have also been shown to activate components of T1R2–T1R3 receptors to induce sweet taste43,47. Although the venus-flytrap domains of both T1R2 and T1R3 are the predominant sites of binding for sucrose, some artificial sweeteners such as cyclamate can act at other sites in the transmembrane domain48. Moreover, some sensory cells, including pancreatic beta cells, can also form T1R3 homodimers that are predicted to be activated at the same domains49. Unravelling such distinctions are necessary to understand the contributions of different organs to the perception of distinctive stimuli.
Conformation change of the sweet taste receptor leads to the activation of a secondary signalling cascade. It begins with the activation of a G protein containing the taste-specific α-subunit, known as α-gustducin. This G protein then activates phospholipase-Cβ2, which uses membrane-anchored phosphatidylinositol 4,5-bisphosphate to catalyze the formation of diacylglycerol and inositol 1,4,5-trisphosphate (IP3). IP3 subsequently activates its receptor to stimulate Ca2+ release from intracellular stores. Increased intracellular Ca2+ then triggers the opening of TRPM4 and TRPM5 channels, calcium-gated non-selective cation channels that allow the intracellular flow of Na+ ions50,51. This depolarization leads to the opening of the voltage-gated non-vesicular ATP CALHM1–CALHM3 channel52, driving the release of ATP from the cell53. Genetic knock out of T1R2–T1R354,55, α-gustducin56, TRPM557 or CALHM1–CALHM352 in mice ablates the sweet taste response in the tongue, confirming the importance of these proteins in sweet taste sensation.
Although first characterized in taste receptor cells of the tongue, T1R2–T1R3 is also expressed in cells of other mucosal surfaces, including the intestines38, bladder58 and lung59. Indeed, taste receptor expression in the intestine has been found on electrically excitable enteroendocrine cells[G]38. Enteroendocrine T1R2–T1R3 activation has been linked to release of the hormones glucagon-like peptide 1 (GLP1) and gastric inhibitory peptide (GIP), which assist in regulating SGLT1 expression60.
단맛 수용체 (Sweet Taste Receptors)
단맛 수용체 서브유닛은
세 개의 독립적인 연구 그룹에 의해 처음 발견되었으며,
주로 혀의 미각 수용체 세포(taste receptor cells) 맥락에서 연구되어 왔다41–43.
앞서 언급한 바와 같이,
단맛은 class C G-단백질 결합 수용체(GPCRs)인
T1R2와 T1R3의 이합체(heterodimer)에 의해 전달된다43,44.
이 GPCRs는
네 가지 주요 도메인을 가지고 있다:
세포외 venus-flytrap 도메인,
시스테인-풍부 linker 도메인,
7회 transmembrane helix 도메인,
그리고 G 단백질과 상호작용하는 세포내 도메인.
다른 수용체와의 상동성(homology)과 모델링에 기반하여,
설탕이 venus-flytrap 도메인에 결합하면
두 flytrap lobe가 닫히는 구조 변화(conformational shift)를 유발하여
세포내 도메인이 활성화되어 G 단백질과 결합한다고 예측된다45,46.
인공감미료도 T1R2–T1R3 수용체의 구성 요소를 활성화하여 단맛을 유발하는 것으로 밝혀졌다43,47.
sucrose의 경우 T1R2와 T1R3의 venus-flytrap 도메인이 주된 결합 부위이지만,
cyclamate와 같은 일부 인공감미료는 transmembrane 도메인의 다른 부위에서 작용할 수 있다48.
또한
췌장 베타세포를 포함한 일부 감각 세포는
T1R3 homodimer를 형성할 수 있으며,
이는 동일한 도메인에서 활성화될 것으로 예측된다49.
이러한 차이를 명확히 밝히는 것은
서로 다른 장기가 독특한 자극에 대한 지각에 기여하는 정도를 이해하는 데 필요하다.
단맛 수용체의 구조 변화는
2차 신호 전달 경로(secondary signalling cascade)를 활성화한다.
이는 taste-specific α-subunit인 α-gustducin을 포함하는 G 단백질의 활성화로 시작된다.
이 G 단백질은
phospholipase-Cβ2를 활성화시키며,
이는 막에 고정된 phosphatidylinositol 4,5-bisphosphate를 이용하여
diacylglycerol과 inositol 1,4,5-trisphosphate (IP3)를 생성한다.
IP3는
이후 자신의 수용체를 활성화하여
세포내 저장소로부터 Ca²⁺ 방출을 자극한다.
증가된 세포내 Ca²⁺은 TRPM4와 TRPM5 채널(칼슘-활성화 비선택적 양이온 채널)을 열어 Na⁺ 이온의 세포내 유입을 허용한다50,51. 이 탈분극(depolarization)은 voltage-gated 비소포성 ATP 채널인 CALHM1–CALHM3을 개방시켜 세포로부터 ATP를 방출한다52,53. T1R2–T1R3 54,55, α-gustducin56, TRPM5 57, 또는 CALHM1–CALHM3 52을 유전자 knockout한 마우스에서는 혀의 단맛 반응이 소실되며, 이는 이러한 단백질들이 단맛 감각에 중요하다는 것을 확인시켜준다.
혀의 미각 수용체 세포에서 처음 특성화되었지만,
T1R2–T1R3은 장(intestines)38, 방광58, 폐59 등 다른 점막 표면의 세포에도 발현된다.
실제로 장에서의 taste receptor 발현은
전기적으로 흥분성인 enteroendocrine cells38에서 발견되었다.
Enteroendocrine T1R2–T1R3 활성화는
SGLT1 발현을 조절하는 데 도움을 주는 호르몬인 glucagon-like peptide 1 (GLP1)과
gastric inhibitory peptide (GIP)의 방출과 연관되어 있다60.
Glucose transporters
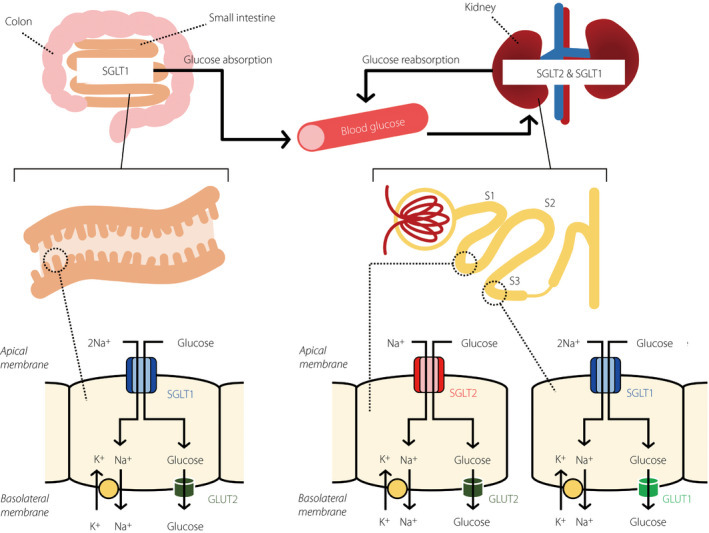
A second class of apical sensors for sugar are glucose transporters. These include the SGLTs and the GLUT family of glucose transporters.
SGLT1 serves as the primary apical transporter for the monosaccharides glucose and galactose, and along with the basal membrane transporter GLUT2, is required for all subsequent post-absorptive glucose signalling61. The protein comprises 14 transmembrane helices and contains a pocket that transports one glucose molecule following two Na+ molecules into the cell62,63. In sensory cells of the gut, this Na+ influx is sufficient to induce depolarization and subsequent opening of voltage-gated L-type calcium channels64. Increased intracellular Ca2+ then triggers the release of peptide hormones such as GLP1 and GIP. Inhibition of SGLT1 pharmacologically or through a genetic knockout showed that this glucose transporter is required for the release of GLP1 and GIP39,65. Recently, it has been demonstrated that intestinal epithelial cells labeled by the cholecystokinin (CCK) promoter release the neurotransmitter glutamate in response to glucose66. This process depends on the activity of SGLT1. Blocking the SGLT1 transporter pharmacologically halts the release of glutamate66. SGLT1 signalling is selective for glucose and is not activated by fructose or artificial sweeteners. Instead, fructose is absorbed by GLUT567. SGLT3 can similarly bind glucose and has been proposed to be a glucose sensor as well68.
Of importance, taste receptor knockout mice show some persistence in sweet taste sensation. This phenomenon suggests the existence of taste receptor-independent mechanisms for detecting sugar in the oral cavity54,55. Recent work has demonstrated that taste receptor cells also express SGLT169,70. Moreover, some of the oral afferent nerve responses to sugar depend on the activity of SGLTs71. Together, these findings suggest that the oral cavity can also utilize SGLT-dependent pathways to sense glucose. Indeed, taste-independent sugar sensing by SGLTs is preserved across species, even in Drosophila72,73.
설탕을 위한 두 번째 apical sensor 클래스는
glucose transporters입니다.
여기에는
SGLTs와 GLUT family의 glucose transporters가 포함됩니다.
SGLT1은
monosaccharides인 glucose와 galactose의 주요 apical transporter로,
basolateral membrane transporter인 GLUT2와 함께
모든 후속 post-absorptive glucose signalling에 필수적입니다61.
이 단백질은
14개의 transmembrane helices로 구성되어 있으며,
두 개의 Na+ 분자가 세포 내로 들어간 후 하나의 glucose 분자를 수송하는 pocket을 가지고 있습니다62,63.
장의 감각 세포에서 이 Na+ 유입은
탈분극(depolarization)을 유발하고,
voltage-gated L-type calcium channels의 개방으로 이어집니다64.
증가된 세포내 Ca2+은
GLP1과 GIP 같은 펩타이드 호르몬의 방출을 유발합니다.
SGLT1을 약리학적으로 억제하거나 유전자 knockout한 연구에서
이 glucose transporter가 GLP1과 GIP 방출에 필수적임이 밝혀졌습니다39,65.
최근 연구에서는
cholecystokinin (CCK) 프로모터로 표시된 장 상피세포가
glucose에 반응하여 신경전달물질 glutamate를 방출한다는 것이 밝혀졌습니다66.
이 과정은
SGLT1의 활성에 의존합니다.
SGLT1 transporter를 약리학적으로 차단하면
glutamate 방출이 중단됩니다66.
SGLT1 signalling은 glucose에 선택적이며,
fructose나 인공감미료에는 활성화되지 않습니다.
대신 fructose는
GLUT5에 의해 흡수됩니다67.
SGLT3도 glucose에 결합할 수 있으며,
glucose sensor로 제안된 바 있습니다68.
중요한 점은
taste receptor knockout 마우스에서도 단맛 감각이 일부 유지된다는 것입니다.
이 현상은
구강 내 설탕 감지를 위한 taste receptor-independent mechanisms의 존재를 시사합니다54,55.
최근 연구에서는
taste receptor cells도 SGLT1을 발현한다는 것이 밝혀졌습니다69,70.
또한
구강 afferent nerve의 sugar에 대한 일부 반응이 SGLTs의 활성에 의존합니다71.
이러한 발견들은
구강에서도 SGLT-dependent pathways를 통해
glucose를 감지할 수 있음을 시사합니다.
실제로
SGLTs에 의한 taste-independent sugar sensing은
Drosophila를 포함한 여러 종에서 보존되어 있습니다72,73.
ATP-gated potassium channels
The majority of the knowledge on KATP channels has been acquired in the context of glucose-dependent insulin secretion in pancreatic beta cells74. KATP channels are also found in oral taste receptor cells and intestinal enteroendocrine cells34,40. Once glucose is transported into a beta cell, it undergoes glycolytic metabolism and culminates in the phosphorylation of ADP to ATP. As the ratio of ATP:ADP increases, KATP channels enter a closed conformational state that depolarizes the cell. This depolarization triggers analogous mechanisms to SGLT1-mediated Ca2+ release, leading to the opening of voltage-dependent L-type calcium channels and subsequent release of vesicles containing peptides or transmitters. This metabolic mechanism is a response to the end products of sugar absorption rather than the sugar molecule itself. As expected, the onset of intracellular metabolic effects of sugar come after the rapid sensing mediated by SGLT1. The existence of both rapid sensing and metabolic responses are in line with the fact that the application of sugar to the intestinal lumen leads to both a millisecond glutamatergic signal and a delayed (~2 min later) hormonal CCK signal66. Activation of the KATP channel is also linked to the release of hormonal peptides such as GLP1 that contribute to regulation of glucose storage and uptake40,64,75.
KATP channels에 대한 대부분의 지식은
췌장 베타세포에서의 glucose-dependent insulin secretion 맥락에서 얻어졌습니다74.
KATP channels는
구강 taste receptor cells과 장 enteroendocrine cells에도 존재합니다34,40.
glucose가 베타세포로 들어가면
glycolysis를 거쳐 ADP가 ATP로 인산화됩니다.
ATP:ADP 비율이 증가함에 따라
KATP channels은 closed conformational state로 들어가 세포를 탈분극시킵니다.
이 탈분극은
SGLT1-mediated Ca2+ 방출과 유사한 메커니즘을 유발하여
voltage-dependent L-type calcium channels을 열고,
펩타이드나 전달물질을 포함한 소포(vesicles)의 방출을 일으킵니다.
이 대사 메커니즘은
설탕 분자 자체가 아니라 sugar absorption의 end products에 대한 반응입니다.
예상대로,
sugar absorption의 intracellular metabolic effects는
SGLT1에 의한 rapid sensing 이후에 나타납니다.
rapid sensing과 metabolic responses의 존재는 장 내강에 sugar를 적용할 때
millisecond 단위의 glutamatergic signal과 (~2분 후) delayed hormonal CCK signal이
모두 발생한다는 사실과 일치합니다66.
KATP channel의 활성화는
GLP1 같은 호르몬 펩타이드 방출과도 연결되어
glucose storage와 uptake 조절에 기여합니다40,64,75.
Intestinal sugar detection and SGLT1
In 2008, de Araujo et al. tested the ability of mice lacking TRPM5 to detect sucrose76. While the mice were unable to distinguish the sweetener sucralose from water, Trpm5 knockout mice retained the ability to distinguish sucrose over water76,77. This finding suggested that sugar preference is independent of sweet taste receptors. Moreover, in intragastric conditioning paradigms, the sweet taste receptor agonist sucralose is insufficient for conditioning a flavour preference78 and both pharmacological and genetic silencing of T1R3 do not alter the reinforcing properties of glucose78.
It became plausible that the ability to recognize sugar depends on a set of receptors and perhaps cells located outside the tongue. Some reports suggested that this receptor could be SGLT1. Its agonist α-methylglucopyranoside can condition a preference, and both pharmacological and genetic silencing of SGLT1 prevent mice from being conditioned to develop a preference for glucose25,79. These results suggest that SGLT1 is the sensor for glucose appetition in the gastrointestinal epithelium.
2008년 de Araujo et al.은 TRPM5이 결핍된 마우스의 sucrose 감지 능력을 테스트했습니다76. 이 마우스들은 인공감미료 sucralose를 물과 구분하지 못했지만, Trpm5 knockout 마우스는 sucrose를 물과 구분하는 능력을 유지했습니다76,77. 이 발견은 sugar preference가 sweet taste receptors와 독립적일 수 있음을 시사했습니다. 또한 intragastric conditioning paradigms에서 sweet taste receptor agonist인 sucralose는 flavour preference conditioning에 충분하지 않았으며78, T1R3의 약리학적·유전적 silencing은 glucose의 reinforcing properties를 변화시키지 않았습니다78.
설탕을 인식하는 능력이 혀 외부에 위치한 receptor 세트와 세포에 의존할 가능성이 제기되었습니다. 일부 보고서는 이 receptor가 SGLT1일 수 있다고 제안했습니다. 그 agonist인 α-methylglucopyranoside는 preference를 conditioning할 수 있으며, SGLT1의 약리학적·유전적 silencing은 마우스가 glucose에 대한 preference를 형성하는 것을 막습니다25,79. 이러한 결과는 SGLT1이 gastrointestinal epithelium에서 glucose appetition의 sensor임을 시사합니다.
Gut sensory transduction
Despite progress in understanding the molecular sensors for sugars, the identity of the gut epithelial cells responsible for the discrimination and subsequent reinforcing properties of sugar remained mysterious for decades4,80,81. Although the connections from gut-to-brain include both humoral and neural pathways82, the prevailing model of nutrient sensation had been that the pre-ingestive senses utilized epithelial transducers synaptically connected to nerves, while post-ingestive senses utilized endocrine cells that secreted hormones83–85.
Studies of sugar sensing in the gut thus emphasized circulating peptides85. These hormonal pathways have mostly been linked to satiety86,87, but some early studies postulated that the desire and preference to consume sugar could be due to the hormone CCK88,89. However, early experiments established that pharmacological inhibition of the CCK A receptor via intraperitoneal injection does not affect preference conditioning for intragastric infusion of the glucose polymer polycose90. In addition, when the hormone peptide YY(3–36) was infused intravenously at low concentrations, food intake decreased and at high concentrations conditioned robust taste aversion91. Hepatic portal vein infusions of GLP1 only reduced food consumption92. The hormone ghrelin is known to stimulate feeding, but pharmacological inhibition and genetic ablation of the ghrelin receptor did not impact sugar conditioning93. Thus, the existing evidence suggests that consumption and preference of sugar is unlikely to rely on gut hormonal signals.
Flavour–nutrient conditioning paradigms showed that the preference and desire to consume sugar depended on sensory cues emanating from the very proximal small intestine. Such an ability of sugar to condition a flavor preference disappears in surgical preparations where the proximal small intestine is bypassed94–96. These studies showed that sugar detection occurred in the duodenum and jejunum, but not in the distal ileum or the stomach94–96. In 2015, it was shown that enteroendocrine cells throughout the intestine and colon of the mouse synapse with nerves97. The discovery of synaptic neurotransmission in these nutrient sensing cells immediately opened the possibility that appetitive decisions and other visceral behaviors could be guided by rapid sensory cues emanating from the gastrointestinal tract98. The endocrine term was limiting for a cell engaging in neurotransmission, so such cells were termed neuropod cells[G]66 (Box 2).
설탕에 대한 molecular sensors에 대한 이해가 진전되었음에도 불구하고,
설탕의 discrimination과 reinforcing properties를 담당하는 gut epithelial cells의 정체는
수십 년 동안 미스터리로 남아 있었습니다4,80,81.
gut-to-brain 연결에는
humoral과 neural pathways가 모두 포함되지만82,
nutrient sensation의 지배적인 모델은
pre-ingestive senses가 nerves와 synaptically 연결된 epithelial transducers를 이용하고,
post-ingestive senses는 호르몬을 분비하는 endocrine cells를 이용한다는 것이었습니다83–85.
따라서
장에서의 sugar sensing 연구는
circulating peptides에 초점을 맞추었습니다85.
이러한 hormonal pathways는
주로 satiety86,87와 연관되어 있지만,
일부 초기 연구에서는 hormone CCK가 sugar consumption과 preference의 원인일 수 있다고 주장했습니다88,89.
그러나
초기 실험에서는 CCK A receptor의 intraperitoneal injection을 통한 pharmacological inhibition이 intragastric infusion of glucose polymer polycose의 preference conditioning에 영향을 미치지 않는 것으로 밝혀졌습니다90. 또한 hormone peptide YY(3–36)을 저농도로 정맥 주입했을 때 food intake가 감소하고, 고농도에서는 robust taste aversion이 conditioning되었습니다91. Hepatic portal vein infusions of GLP1은 food consumption만 감소시켰습니다92. Ghrelin은 feeding을 자극하는 것으로 알려져 있지만, ghrelin receptor의 pharmacological inhibition과 genetic ablation은 sugar conditioning에 영향을 미치지 않았습니다93. 따라서 기존 증거는 sugar consumption과 preference가 gut hormonal signals에 의존할 가능성이 낮다는 것을 시사합니다.
Flavour–nutrient conditioning paradigms는 sugar의 preference와 desire to consume가 proximal small intestine에서 나오는 sensory cues에 의존한다는 것을 보여주었습니다. proximal small intestine을 bypass하는 surgical preparations에서는 sugar의 flavour preference conditioning 능력이 사라집니다94–96. 이러한 연구들은 sugar detection이 duodenum과 jejunum에서 발생하며, distal ileum이나 stomach에서는 발생하지 않는다는 것을 보여주었습니다94–96. 2015년, 마우스의 장과 colon 전체에 enteroendocrine cells가 nerves와 synapse를 형성한다는 것이 밝혀졌습니다97. 이러한 nutrient sensing cells에서의 synaptic neurotransmission 발견은 gastrointestinal tract에서 나오는 rapid sensory cues가 appetitive decisions와 다른 visceral behaviors를 유도할 가능성을 즉시 열었습니다98. endocrine term은 neurotransmission에 참여하는 세포에게 제한적이었기 때문에, 이러한 세포를 neuropod cells66라고 명명했습니다 (Box 2).
Box 2 |. Neurotransmission from gut to brain.
In 2011, an anatomical peculiarity was observed in a subset of enteroendocrine cells that opened a new mechanism for intestinal communication. Some of these sensory cells had a cytoplasmic protrusion at the base of the cells which resembled a neuronal axon171. Subsequent work confirmed that some enteroendocrine cells, both with and without a basal extension, could from synapses with enteric, vagal and splanchnic nerves66,97,172,173. To account for the neuron-like function in these cells, the term neuropod cell was extended to those epithelial cells engaging in neurotransmission66. This gut neuroepithelial sensory circuit could serve to account for the chemical, mechanical and thermal properties of ingested nutrients. The similarity between neuroepithelial circuits in the gastrointestinal tract and other sensory systems like those of taste, olfaction and vision raised the hypothesis that there exists continuity between pre-ingestive and post-ingestive sensory mechanisms.
Neuropod cells are a subset of enteroendocrine cells that are differentiated by their neural function. Transcript and protein analyses have shown that subpopulations of enteroendocrine cells express pre-synaptic proteins such as synapsin 1, piccolo, bassoon, MUNC13B, RIMS2, latrophilin 1 and neurexin 297. Ultrastructural investigations have revealed that neuropods contain neurofilaments (see the figure, part a) as well as both dense core vesicles containing peptides and synaptic clear vesicles containing neurotransmitters (see the figure, part b)164. In vitro, neuropod cells synapse with co-cultured neurons, and optogenetic activation of neuropod cells triggers excitatory postsynaptic potentials in connected neurons within 60 milliseconds66. In vivo, monosynaptic tracing using rabies virus demonstrated that small intestinal neuropod cells form monosynaptic connections with vagal neurons to rapidly communicate nutrient sensory information to the brain (see the figure, part c)66. In a series of studies utilizing organoid cultures, Bellono et al. showed that colonic enterochromaffin cells synapse with peripheral nerves to rapidly transduce the presence of irritants using serotonin172. Subsequent results showed that these cells could also form purinergic synapses173. In vivo, luminal optogenetic manipulation of neuropod cells shows that these cells are required for sugar preference99. Parts a and b adapted with permission from REF.164. Part c adapted with permission from REF.66.
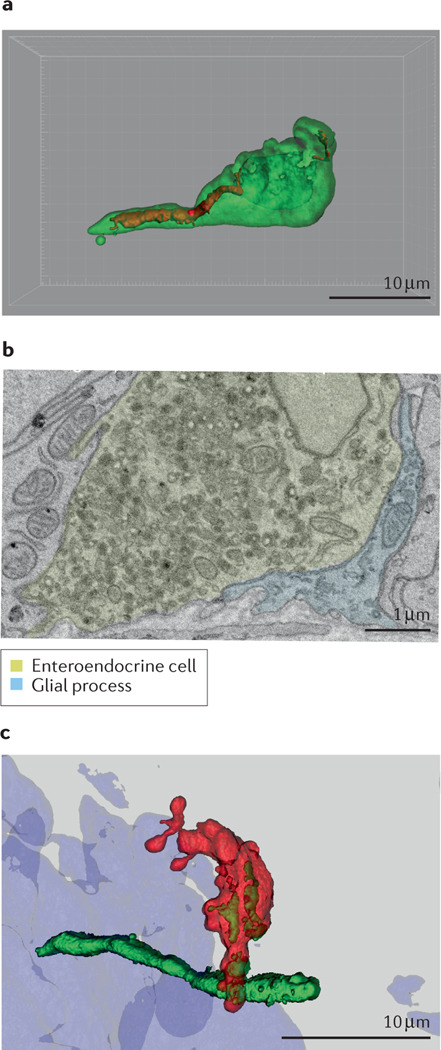
In 2018, Kaelberer et al. showed that duodenal neuropod cells synapse with vagal neurons to convey the presence of sugar66. These cells express SGLT1 and increase intracellular Ca2+ in response to glucose. Glucose perfusion onto these cells induces the release of glutamate, which can evoke excitatory postsynaptic currents in co-cultured neurons66. Silencing duodenal neuropod cells using optogenetics or pharmacological inhibitors of ionotropic glutamatergic receptors completely abolished the rapid phase of vagal firing induced by glucose infusion. Although these cells were first identified using the CCK promoter, rapid transmission of sugar stimuli onto vagal afferents is independent of the hormone CCK. Inhibition of the CCKA receptor with the pharmacological inhibitor devazepide does not affect rapid vagal firing induced by glucose infusion in the duodenum. Thus, these findings established that the ability of the brain to receive in milliseconds sensory stimuli from sugar entering the intestine depends on glutamatergic signaling from neuropod cells.
In 2022, Buchanan et al. developed a new tool to interrogate the contribution of visceral sensory cues to behaviour. They developed a flexible fibre optic cable to drive light-sensitive opsins in specific sensory cells exposed to the gut lumen. In this way, it was now possible to interrogate the contribution of neuropod cells to sugar preferences in real time99. When trained mice are presented with two bottles containing either sucralose or isosweet sucrose solutions, they prefer sucrose. However, silencing duodenal neuropod cells labeled by the CCK promoter prevents the animals from distinguishing sugar from artificial sweetener. Moreover, pharmacological inhibition of ionotropic glutamate receptors in the duodenum also extinguished preference for sugar over sweetener. These findings demonstrated that the animal distinguishes sugar from sweetener guided by intestinal glutamatergic signalling. In other words, neuropod cells guide the animal’s consumption and preference for sugar by rapidly detecting sugar. Whether glutamatergic neurotransmission from neuropod cells is necessary for other behaviours such as the development or recall of sugar preferences remains to be tested.
From the gut to the brainVagal pathways
The vagus nerve contains both sensory and motor fibres that connect the brainstem with the viscera, including the gut100,101. The entry of nutrients into the duodenum stimulates vagal firing102, and specific vagal neurons are tuned to macronutrients103. Both sugars66 and artificial sweeteners99 entering the duodenal lumen rapidly stimulate vagal firing, but the vagal responses appear to be mediated by specific subpopulations of sensory vagal nodose neurons (Fig. 2)81,104. Vagal responses to sucrose depend on SGLT181 and are confined to the small intestine as colonic perfusion of sucrose does not increase vagal firing99. Although the colonic mucosa is sparsely innervated by the vagus nerve, SGLT1 expression is absent in the colonic epithelium99. Importantly, vagal nerve terminals do not reach the lumen of the intestine105,106 and do not express SGLT199,105,107. Therefore, vagal neurons must engage with intestinal neuropod cells that act as the primary sensory transducers. And at least for sugars, vagal neurons depend on sensory inputs from neuropod cells. Whether the vagus serves as a relay from the intestine to brain or has some role in integrating information from multiple primary sensors in other visceral organs remains to be tested.
Fig. 2 |. Heterogeneity within nodose neurons.
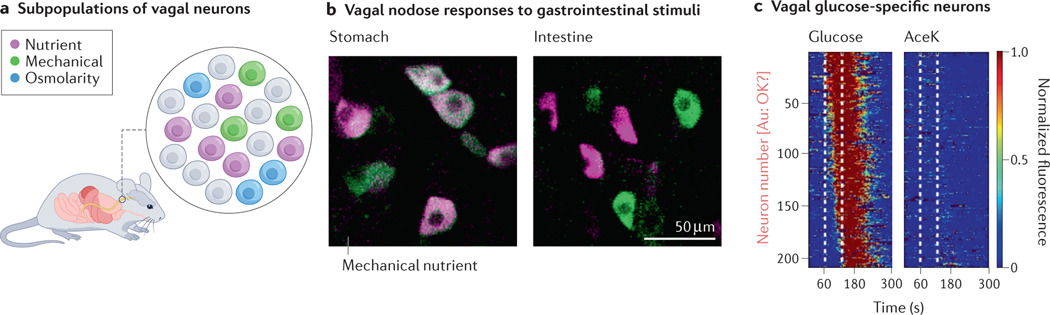
a | Schematic showing individual subpopulations of vagal sensory nodose neurons that respond to different stimuli. b | GCaMP3 calcium responses of nodose neurons in response to mechanical stretch and nutrient perfusion. In the stomach, most activated neurons are activated by stretch, while in the intestine, distinct populations respond to mechanical and chemical stimuli. c | GCaMP6f imaging of vagal nodose neurons. Subpopulations of neurons imaged were selective for glucose and not artificial sweetener acesulfame K (AceK). Only neurons that had a response to glucose are shown. Each row represents an individual cell, and colour represents normalized fluorescence. Part b adapted with permission from REF.104. Part c adapted with permission from REF.81.
Despite its central role in gut–brain signaling, the vagus nerve’s function in sugar preference has been debated. For over a century, the prevailing idea was that vagal signalling from the small intestine to the brain is a negative feedback signal to inhibit future consumption4,108. In 2018, Han et al. countered this theory by identifying a population of “reward neurons” in the right vagus109. Using optogenetics together with viral tracing, they manipulated a specific population of vagal afferents innervating the proximal gut independently of efferents or fibres that do not innervate the gut. Activating these gut-innervating neurons was sufficient for mice to perform arbitrary operant behaviours such as nose poking. Moreover, stimulating these same neurons was sufficient to condition a preference for an arbitrary location. Such self-stimulating behaviours confirmed the existence of vagal afferents capable of inducing reward109. The study went on to use these vagal neurons to modulate the rewarding effects of intragastric perfusions of fat emulsion, and future work should address whether sugars engage similar circuits.
The challenge of investigating the function of the vagus lies in a lack of tools to target specific subpopulations of vagal afferents110. Classic techniques such as surgical vagotomy are limited for several reasons. The vagus nerve can regrow111,112, nerve transection does not fully differentiate between afferents and efferents, and nerve transection is often not organ specific. Such limitations produce results confounded by many unintended effects such as high mortality rate in experimental animals, failure to thrive, change in gastric motility and emptying, and other secondary effects. Pharmacological approaches such as capsaicin ablation are also incomplete. Nevertheless, these techniques have been used in numerous studies to conclude that reward learning from intra-intestinal glucose is vagal-independent96,113–115.
A recent study used a genetic tetanus toxin strategy in mice to silence all sensory vagal neurons expressing the vesicular glutamate transporter 2 in the nodose ganglion[G]. The study found that these mice did not prefer glucose over an artificial sweetener81. Such seemingly contradictory pieces of evidence may point to redundancy in the system. When stimuli are well separated in time, other signalling pathways, including humoral and post-absorptive hepatic signalling, may be recruited to assure proper reinforcement and association116,117. However, in complex scenarios in which multiple stimuli are consumed within one meal, gut–neural circuits may be engaged to ensure proper stimulus discrimination and association118.
New efforts to subclassify vagal neurons guided by transcription patterns have identified that vagal neurons innervating the mucosa of the proximal small intestine specifically express genes such as Gpr65 or Ntsr1105,107. Cre-lines to specifically modulate these subpopulations have also been developed, and such tools will enable future experimentation to differentiate the function of specific vagal subpopulations104,119.
Splanchnic pathways
Another set of neurons that project from the intestine to the brain are splanchnic nerves. These nerves derive from the celiac and superior mesenteric ganglia and enter the brain via the spinal cord. Limited evidence exists to determine the function of these neurons in sugar sensing. While some studies have not observed significant changes in food consumption120,121, surgical ablation of these afferents using a complete transection of the ganglia reduces the strength of conditioned preferences for glucose115. More recently, splanchnic lesions were shown to reduce the responsiveness of central feeding circuits in response to glucose122. Similar to vagotomies, surgical transection of splanchnic nerves to interrogate their contribution to behaviour has several limitations. Future intersectional genetic approaches using viruses carrying chemical or optical actuators will help elucidate the contribution of these nerves to rapid appetitive decisions.
Central circuits in sugar preference
Neuronal signals from the viscera enter the brain through the brainstem before they are conveyed to a distributed network of overlapping circuits throughout the brain involved in controlling behaviour. Here, we describe the primary brainstem neurons that integrate signals first encountered by the vagus nerve, the current understanding of the role of upstream dopaminergic reward circuits in the basal ganglia, and homeostatic feeding circuits in the hypothalamus involved in triggering consumption and learning the value of sugar.
Nucleus tractus solitarius
Afferent terminals from pre-absorptive circuits terminate in the nucleus tractus solitarius (NTS). This nucleus forms a column of cell bodies within the medulla oblongota with general topographic distribution depending on the location of the input123. The rostral portion of the NTS (rNTS) receives inputs mainly from the oral cavity, while the more caudal portion (cNTS) receives glutamatergic inputs from visceral organs, including the gut106,124. NTS neurons also receive inputs from humoral circulating factors such as leptin, as well as feedback from neural circuits arising in the hypothalamus125,126. NTS neurons project to many regions across the brain, including the dorsal motor nucleus of the vagus, hypothalamus, central nucleus of the amygdala, parabrachial nucleus, and other brainstem areas involved in adrenergic (locus coeruleus), dopaminergic (ventral tegmental area) and serotonergic signalling (dorsal raphe)127,128. NTS neurons are primarily glutamatergic and co-release peptides such as norepinephrine, GLP1 and CCK129. The NTS forms a heterogeneous population of neurons that can induce both satiation and appetition130,131.
Distinct populations of cNTS neurons, as measured by cFos expression, are activated by intragastric and intestinal glucose perfusions132,133. Tan et al. recently identified that a population of proenkephalin-expressing neurons within the cNTS is activated by duodenal glucose infusion81. These same neurons are not activated by the artificial sweetener acesulfame K. Acesulfame K binds to sweet taste receptor T1R2–T1R3 but does not serve as a substrate of the sugar receptor SGLT1. Ablating synaptic activity in these brainstem neurons using tetanus toxin does not prevent discrimination of either glucose or the artificial sweetener from water. However, their manipulation does prevent mice from discriminating between glucose and artificial sweetener, and from subsequently developing a preference for glucose over sweetener in a 48-hour test. Moreover, chemogenetically activating these neurons was sufficient to condition a flavour preference81. Monosynaptic tracing studies from neuropod cells in the duodenum have shown connections with vagal afferent neurons that project to the NTS66. These vagal neurons most likely overlap with the peripheral reward neurons that project to NTS proenkephalin-expressing neurons, but this hypothesis remains to be tested.
Dopaminergic reward circuits
Reward and reinforcement learning are linked to dopaminergic circuits that facilitate goal-directed behaviours and promote the ‘wanting’ of food rewards134,135. These circuits are conceptualized to mediate subconscious aspects of food reward and selection4. Eliminating dopaminergic signalling in mice does not prevent sugar consumption, and mice can still discriminate sugar from water136. This deficiency did reduce the total number of consumption bouts[G], but the rate of licking and bout size were greater136, suggesting that dopamine is not required to detect sucrose palatability[G]. By contrast, pharmacological inhibition of dopamine D1 receptors in the striatum during training sessions but not during testing sessions prevented flavour preference conditioning137. Thus, dopamine signalling underlies the learned associations between food value and environmental cues rather than intrinsic sugar preferences.
Two independent dopaminergic circuits arise in the midbrain: the mesolimbic[G] system projecting from the ventral tegmental area (VTA) to the ventral striatum, and the nigrostriatal[G] system projecting from the substantia nigra pars compacta (SNPc) to the dorsal striatum (Fig. 3). Outputs from these two basal ganglia regions project to motor feeding circuits to control behaviour. Oral and intra-intestinal infusion of sucrose shows increased activation of neurons and dopamine release in both pathways77,138–141. However, in sham-fed rats, in which the oral cavity is exclusively stimulated, sucrose only induces dopamine release in the ventral striatum139. Thus, while the mesolimbic system receives signals from oral and post-ingestive sites, the nigrostriatal system only receives input from the intestine and post-absorptive sensors.
Fig. 3. |. Dopaminergic circuits in sugar sensing.
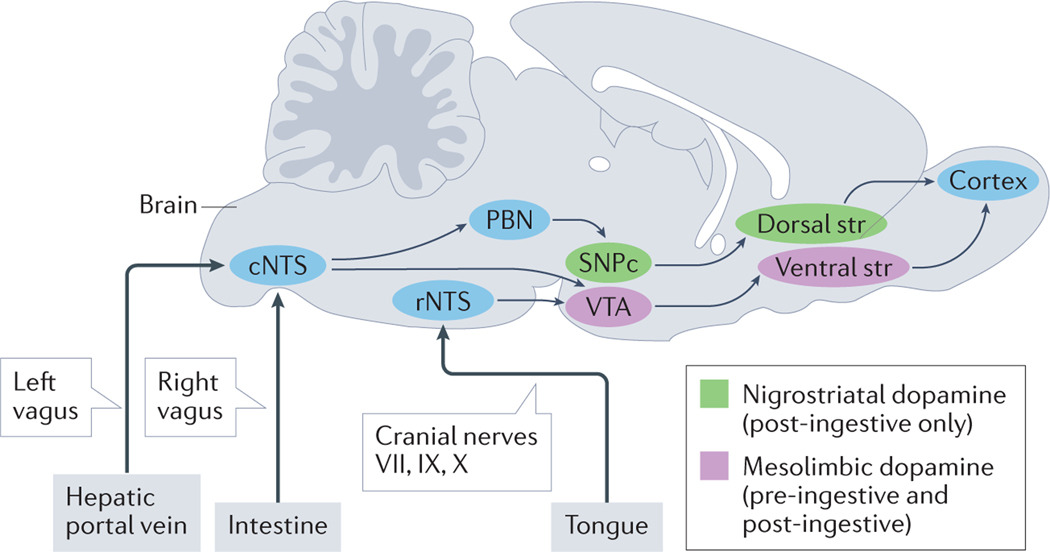
Two distinct dopaminergic pathways are activated by the experience of sugar in the mouse brain. Mesolimbic dopamine release triggered by connections between the ventral tegmental area (VTA) and the ventral striatum (Str) are produced in response to oral, intestinal and hepatic portal vein infusion of sugar. By contrast, nigrostriatal dopamine release between the substantia nigra pars compacta (SNPc) and the dorsal striatum are only triggered by hepatic portal vein and intestinal signalling in response to sugar. All peripheral inputs enter the brain through the nucleus tractus solitarius (NTS), where oral inputs enter the rostral (rNTS) portion, and intestinal and post-absorptive signals enter via the caudal (cNTS) portion. The NTS can then project to dopaminergic circuits directly or via polysynaptic circuits including the parabrachial nucleus (PBN).
To compare the functions of the mesolimbic and nigrostriatal pathways in intestinal sensing, Tellez et al. measured dopamine release in both the ventral and dorsal striatum while mice licked sucralose and received intragastric infusions of either sucralose or glucose141. While both solutions induced dopamine release in the ventral striatum, only intragastric glucose induced dopamine release in the dorsal striatum. Suppressing taste-mediated signalling by mixing in a bitter tastant reduced ventral striatal dopamine release. And suppressing gut-mediated signalling using a sugar analogue that cannot be absorbed reduced dorsal striatal release. In addition, using both cell ablation and optogenetic activation of neurons expressing dopamine D1 receptors in the dorsal and ventral striatum, it was determined that the nigrostriatal pathway is both necessary and sufficient for the post-ingestive reinforcing effect of glucose141. This study demonstrates independent roles for the mesolimbic and nigrostriatal dopamine pathways in learning the value of consuming glucose. Their role in the immediate acceptance of glucose is less clear, and the relative contributions of intestinal and post-absorptive signalling on nigrostriatal dopamine release remains to be established.
Activation of mesolimbic dopamine neurons using optogenetics is sufficient to drive voluntary ingestion of water142. Moreover, in two-bottle preference tests designed to test the ‘reward value’ of dopaminergic signalling, mesolimbic activation was more rewarding than non-caloric sweetener sucralose but less rewarding than sucrose142. In this regard, mice can reverse their natural preference for sucrose over sucralose if sucralose is paired with mesolimbic activation142. Thus, the mesolimbic system assigns more value to solutions than solely taste receptor-mediated mechanisms but less value than SGLT1 and taste receptor mechanisms together.
Recently, miniscopes implanted in the brain were used to directly image individual VTA neuron activity in freely behaving mice. Intra-gastric infusion of sucrose positively modulated a greater proportion of dopaminergic neurons in the VTA than did intra-gastric infusion of sucralose143. It was also shown that burst firing of VTA dopaminergic neurons is required for the reinforcing effects of intra-gastric sucrose in a food-seeking task. Transection of the hepatic vagus nerve blunted VTA responses, suggesting that the mesolimbic activation depends on post-absorptive signalling143. These results suggest that midbrain neurons, specifically those mesolimbic dopaminergic neurons, assign the reward value of sugars independently of their sweet taste.
Midbrain dopaminergic centres receive inputs from NTS neurons that project directly or via the lateral parabrachial nucleus129. In tracing experiments, rNTS neurons polysynaptically target the VTA, while cNTS neurons project monosynaptically or polysynaptically to the VTA109,144 and polysynaptically to the SNPc109. Viral tracing studies also confirmed that the nigrostriatal pathway receives polysynaptic input from the reward neurons of the right vagus109. Activation of right vagal reward neurons increased dopamine release in the dorsal striatum but not the ventral striatum109. Parallel studies showed that optogenetic activation of the left vagus increased activity of VTA dopaminergic neurons143. These results imply an asymmetry within the vagus, with left vagal stimulation increasing mesolimbic activation and right vagal stimulation increasing nigrostriatal activation.
Agouti-related peptide feeding circuits
Hypothalamic neurons have long been known to be activated by intra-intestinal infusions of nutrients, including glucose145. Subsequent studies identified specific neuronal populations involved in controlling food intake, including the agouti-related peptide (AgRP) neurons of the arcuate nucleus146,147. Activity of AgRP neurons is strongly correlated with the energy state of the animal148, and stimulation of these neurons with optogenetics or chemogenetics leads to appetitive behaviours, including food consumption and seeking149,150. This consumption is dependent on the nutrient composition of the food consumed, as AgRP neuron activation leads to rapid consumption of sucrose but not saccharin pellets151. By contrast, ablation of these neurons leads to a loss of desire to consume food and failure to thrive147. AgRP neurons project downstream to several brain areas involved in food consumption, such as the lateral hypothalamus, paraventricular hypothalamus and the bed nucleus of the stria terminalis152.
Although AgRP neurons receive synaptic inputs from diverse brain areas including the cNTS, they are also adjacent to the median eminence and thus were traditionally thought to respond to circulating humoral factors153. Recent direct imaging of AgRP calcium activity in awake mice has shown that AgRP neurons are rapidly and strongly suppressed by nutrient consumption154–156. The timing of the response was more rapid than possible by humoral mechanisms, suggesting that AgRP neurons may receive information via polysynaptic pathways from the viscera. The rapid suppression of AgRP neuron activity occurs in response to intragastric infusion of glucose and not sucralose, mirroring the behavioural studies, and further confirming the importance of nutrient identity156. AgRP suppression by intraduodenal infusion of glucose depends on SGLT1122. Moreover, the responses are proportional to the caloric load of glucose and are thought to represent predicted nutrient consumption155.
Given the suppression of AgRP neurons in response to nutrients, it was postulated that AgRP activation could provide an instructive negative valence signal to prevent future consumption. Concordantly, continuous stimulation of AgRP neurons was sufficient to condition flavour and place avoidances154. By contrast, the rapid suppression of AgRP neuron activity, as seen with physiological infusion of sugar, was shown to have a positive valence and was sufficient to condition flavour-nutrient preferences157. In addition, AgRP neuron activation potentiates the dopamine release in the nucleus accumbens and VTA dopaminergic neuron activity in response to food stimuli158,159. Notably, AgRP responses can also occur to visual, olfactory and conditioned cues that precede nutrient delivery, suggesting that one of the functions of AgRP neurons is to predict imminent nutrient availability155,160. These responses diminish rapidly without continual intraintestinal stimulation154.
The identity of the visceral sensors and inputs for glucose-mediated AgRP suppression remains unclear. Recent work suggests that AgRP suppression is unaffected by vagotomy and is reduced by transection of splanchnic nerves122. Moreover, several studies have implicated the role of peptides such as CCK and PYY in reducing AgRP activity155,156. However, the signal for glucose-mediated changes is still unidentified, and the neural route from the intestine or post-absorptive sites to the arcuate nucleus is an area of active research105.
Integrated view
The gut’s ability to detect, distinguish and transduce stimuli from individual nutrients like glucose is a fundamental sense. This sense is critical for animals to recognize the contents that they are actively transporting from the external environment into their internal milieu. The current evidence thus leads us to propose a unified model of detecting sugar that involves continuous sensing along the pre-absorptive epithelium (Fig. 4).
Fig. 4 |. A model of neuroepithelial circuits for sugar sensing.
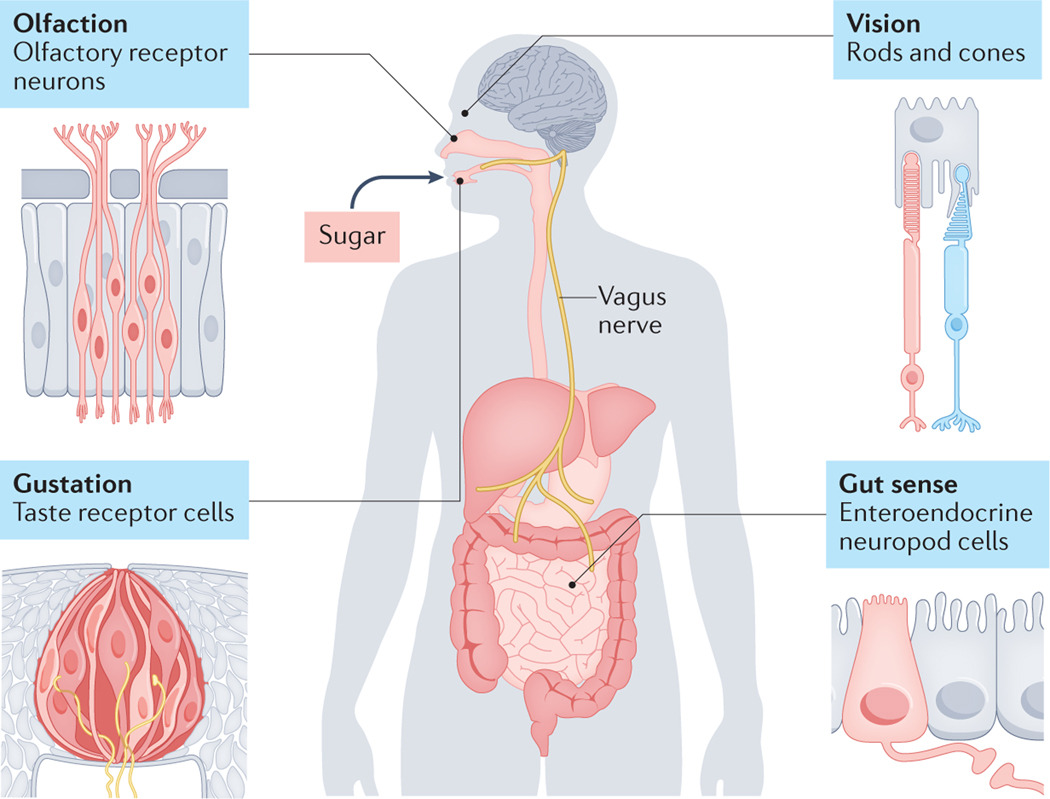
Chemosensation through synaptic neural circuits occurs via rods and cones in the eye, olfactory receptor neurons in the nose, taste receptor cells in the tongue, and neuropod cells in the intestine. This forms a continuous sensory system that monitors sugar from the process of its foraging to absorption.
In this model, the sensory experience of sugar begins before it is even ingested. Visual, olfactory and even tactile cues trigger central feeding circuits that predict the value of its ingestion. Once it enters the oral cavity, it can activate taste receptor cells that signal through cranial nerves and the rNTS into the gustatory cortex for conscious perception as well as into mesolimbic dopaminergic circuits for hedonic[G] value.
As sugar is ingested and contacts the stomach and proximal duodenum, neuropod cells sense it through SGLT1, and release glutamate onto vagal afferents that project to the caudal NTS, probably onto proenkephalin-expressing neurons. These neurons send projections to the nigrostriatal dopamine system, as well as the mesolimbic dopamine system and also likely engage rapid suppression of AgRP neuron activity. Simultaneously, other enteroendocrine cells begin releasing humoral and paracrine factors like GIP, GLP1 and CCK.
Finally, enterocytes actively absorb sugar via SGLT1 and deposit it into the portal venous circulation, where hepatic sensors can become activated and signal the absorbed quantity of glucose to the CNS via circulating factors and left vagal activation. The combination of intestinal epithelial signalling and portal venous signalling converge on higher-order dopaminergic and homeostatic circuits to drive acceptance of the solution and subsequent reinforcement learning.
Recognizing the combined role of the entire sensory system will probably improve future efforts to combat dietary imbalances. Artificial sweeteners cannot improve dietary choices alone because while artificial sweeteners can trick oral taste-receptor signalling, they do not trigger the intestinal SGLT1-dependent glutamatergic signalling pathway. Gut sensations are also increasingly linked to other important nervous system functions. Arousal circuits and sleep patterns can be disrupted by visceral sensation161. Emotional states can be modulated by intestinal microbial and nutrient stimuli162. Even memories formed in the hippocampus can be altered by intestinal signalling163. Each of these functions involves the gut sense. Unraveling these manifold abilities will allow us to harness the gut to enjoy pleasurable foods, foster healthy habits and promote emotional well-being. After all, the instinctive wisdom of the body is the reason why, when it relates to sugar, we go with our gut.
Acknowledgements
The authors thank S. Simon, R. Gutierrez, I. de Araujo, and A. Sclafani for their early review of the manuscript. They also thank E. Bohórquez, M. Kaelberer, L. Rupprecht and P. Weng for their helpful edits.
GLOSSARYArtificial sweeteners
Chemicals produced to mimic sugar by binding to the sweet taste receptor while also not being capable of being absorbed
Exteroceptive
Sensitivity to stimuli originating outside the epithelial barrier of the body
Interoceptive
Sensitivity to stimuli originating inside the epithelial barrier of the body
Satiation
A process that suppresses nutrient intake and stops current consumption
Satiety
A sense of fullness that persists after eating that suppresses future consumption
Reinforcement
A procedure that results in the frequency or probability of a response being increased in such a way
Appetition
A process that promotes nutrient intake
Operant
A response that produces a consequential effect on the environment
Flavour–nutrient conditioning
A form of classical conditioning where flavours of a solution are paired to infusions of chemical stimuli into the gut
Sweet
The perception of taste associated with sugar, typically triggered by activation of sweet taste receptors in the tongue
Enteroendocrine cell
Sensory epithelial cell of the gut that releases hormones in response to various stimuli
Neuropod cells
Sensory epithelial cells of the gut that synapse with nerves
Nodose ganglion
The sensory ganglion of the vagus nerve located in the jugular foramen
Bouts
Instances of food consumption
Palatability
The degree of pleasantness or reward associated with stimulating the oral palate
Mesolimbic system
A network of dopaminergic neurons consisting of the ventral striatum that receives input from the ventral tegmental area and is related to emotion and reward
Nigrostriatal system
A network of dopaminergic neurons consisting of the dorsal striatum that receives input from the substantia nigra pars compacta and is linked to action initiation
Hedonic
A degree of pleasantness or reward induced by an interaction or thought
Footnotes
Competing interests
The authors declare no competing interests.
References
- 1.Lutter M. & Nestler EJ Homeostatic and hedonic signals interact in the regulation of food intake. J. Nutr. 139, 629–632 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Rossi MA & Stuber GD Overlapping brain circuits for homeostatic and hedonic feeding. Cell Metab. 27, 42–56 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Yudkin J. Pure, White, and Deadly: How Sugar Is Killing Us and What We Can Do to Stop It. (Penguin, 2013). [Google Scholar]
- 4.De Araujo IE, Schatzker M. & Small DM Rethinking food reward. Annu. Rev. Psychol. 71, 24.1–24.26 (2020). [DOI] [PubMed] [Google Scholar]
- 5.Zuker CS Food for the brain. Cell 161, 9–11 (2015). [DOI] [PubMed] [Google Scholar]