
- 인간 단맛 수용체(T1R2/T1R3 heterodimer)의 고해상도 구조와 기능을 처음으로 상세히 밝힌 연구.
- Cryo-EM 등 첨단 구조생물학 기법을 활용해 수용체의 3차원 구조, ligand (설탕·인공감미료) 결합 부위, 활성화 메커니즘을 규명.
주요 발견
- T1R2와 T1R3의 venus flytrap domain, transmembrane domain 등 주요 구조 도메인 상세 분석.
- 다양한 단맛 물질(자연 설탕, 인공감미료)이 결합하는 방식과 receptor 활성화 과정의 차이 규명.
- 구조 기반으로 단맛 강도·특이성을 설명하고, 새로운 감미료 개발이나 수용체 조절 약물 설계의 기초 제공.
의의
- 단맛 감각의 분자적 이해를 크게 진전시킴.
- 비만·당뇨·식욕 조절 연구, 그리고 저칼로리 감미료 개발에 중요한 구조적 근거 제공.
한 줄 요약: 인간 단맛 수용체(T1R2/T1R3)의 고해상도 구조와 기능적 특성을 밝힌 2025년 Nature 연구
Abstract
Sweet taste perception influences dietary choices and metabolic health. The human sweet taste receptor, a class C G-protein-coupled receptor (GPCR) heterodimer composed of TAS1R2 and TAS1R3 (refs. 1,2), senses a wide range of sweet compounds—including natural sugars, artificial sweeteners and sweet proteins—and affects metabolic regulation beyond taste. However, the lack of three-dimensional structures hinders our understanding of its precise working mechanism. Here we present cryo-electron microscopy structures of the full-length human sweet taste receptor in apo and sucralose-bound states. These structures reveal a distinct asymmetric heterodimer architecture, with sucralose binding exclusively to the Venus flytrap domain of TAS1R2. Combining mutagenesis and molecular dynamics simulations, this work delineates the sweetener-recognition modes in TAS1R2. Structural comparisons further uncover conformational changes upon ligand binding and a unique activation mechanism. These findings illuminate the signal transduction mechanisms of chemosensory receptors in the class C GPCR family and provide the molecular basis for the design of a new generation of sweeteners
단맛 인지는
식이 선택과 대사 건강에 영향을 미친다.
인간 단맛 수용체는 T
AS1R2와 TAS1R3로 구성된 class C
G-단백질 결합 수용체(GPCR) 이합체로1,2,
천연 설탕, 인공감미료, 단백질성 단맛 물질 등 다양한 단맛 화합물을 감지하며,
미각을 넘어 대사 조절에도 영향을 미친다.
그러나
3차원 구조의 부재로 인해 정확한 작동 메커니즘에 대한 이해가 제한되어 왔다.
본 연구에서는
apo 상태와 sucralose 결합 상태의 full-length 인간 단맛 수용체에 대한
cryo-electron microscopy 구조를 제시한다.
이 구조들은
TAS1R2의 Venus flytrap domain에 sucralose가 독점적으로 결합하는
독특한 비대칭 이합체 구조를 드러낸다.
돌연변이 분석과 분자동역학 시뮬레이션을 결합하여
TAS1R2에서의 감미료 인식 모드를 상세히 규명하였다.
구조 비교를 통해
리간드 결합 시 일어나는 conformational change와 독특한 활성화 메커니즘을 밝혀냈다.
이러한 발견은 class C
GPCR 계열 화학감각 수용체의 신호 전달 메커니즘을 밝히고,
차세대 감미료 설계를 위한 분자적 기초를 제공한다.
Taste is a fundamental sensory modality that has an important role in human life, guiding food choice and dietary behaviours. Humans perceive five basic taste stimuli: sweet, bitter, sour, salty and umami3–6 , with sweet taste being essential for energy homeostasis. Three of these taste modalities—sweet, umami and bitter—are mediated by GPCRs7 . Specifically, sweet and umami tastes are detected by type 1 taste receptors (TAS1Rs)8,9 , which are class C GPCRs, whereas bitter tastes are sensed by type 2 taste receptors (TAS2Rs)10–12, which are categorized as class T GPCRs. Both receptor types are expressed in specialized taste receptor cells (TRCs) in the taste bud and signal primarily through the Gα subunit of gustducin (Gαgust), initiating downstream physiological responses13,14. The TAS1Rs include three members—TAS1R1, TAS1R2 and TAS1R3 (refs. 8,9)—that form heterodimers to mediate sweet (TAS1R2–TAS1R3 heterodimer) or umami (TAS1R1–TAS1R3 heterodimer) taste perception1,2,15,16 (Fig. 1a). In addition to TAS1Rs, the class C GPCR family also includes metabotropic glutamate receptors (mGluRs)17, γ-aminobutyric acid receptors (GABABRs)18, calcium-sensing receptor (CaSR)19 and several orphan receptors20. TAS1Rs share the characteristic of class C GPCRs, which consist of an extracellular venus flytrap domain (VFTD), a cysteine-rich domain (CRD) and the seven-transmembrane domain (TMD). The human sweet taste receptor recognizes diverse types of sweettasting resources, including natural sugars such as glucose and sucrose, artificial sweeteners such as sucralose and neotame21, and naturally occurring sweet proteins such as monellin, thaumatin, miraculin and brazzein22,23. Of note, the receptor exhibits low binding affinity for sugars, typically in the micromolar to millimolar range24,25 (Fig. 1b). Sweet taste perception is closely linked to energy metabolism because it serves as an early and crucial sensory signal that guides organisms towards energy-rich foods26,27. Beyond oral taste perception, the TAS1R2 and TAS1R3 receptors are ectopically expressed in the gut28, pancreas29 and brain30, where they are involved in regulating metabolic processes. Therefore, the ectopic TAS1Rs have emerged as potential therapeutic targets for obesity and metabolic disorders, including diabetes and hyperlipidemia29,31,32. Despite its chemosensory and physiological importance, the structural characterization of the human sweet taste receptor remains a major challenge. Previous structural studies have only reported the crystal structures of the VFTD in TAS1Rs from medaka fish bound to amino acids33. However, the full-length structures of human TAS1Rs remain unknown, limiting our understanding of sweetener modulation and receptor activation mechanisms. Here we present cryo-electron microscopy (cryo-EM) structures of the full-length human TAS1R2– TAS1R3 heterodimer in both apo and sucralose-bound states, providing a structural template for understanding sweet taste perception at molecular level.
미각은
인간의 삶에서 중요한 역할을 하는 기본적인 감각 양식으로,
음식 선택과 식이 행동을 안내한다.
인간은
다섯 가지 기본 미각 자극—단맛, 쓴맛, 신맛, 짠맛, 감칠맛3–6—을 인지하며,
그중 단맛은 에너지 항상성(energy homeostasis)에 필수적이다.
이 중 단맛, 감칠맛, 쓴맛은
G-단백질 결합 수용체(GPCRs)에 의해 매개된다7.
특히 단맛과 감칠맛은
type 1 taste receptors (TAS1Rs)8,9에 의해 감지되며,
이는 class C GPCRs에 속하고,
쓴맛은 type 2 taste receptors (TAS2Rs)10–12에 의해 감지되며 class T GPCRs로 분류된다.
두 수용체 유형 모두
맛봉오리(taste bud) 내 특화된 taste receptor cells (TRCs)에 발현되어 있으며,
주로 gustducin의 Gα subunit (Gαgust)를 통해 신호를 전달하여
하위 생리적 반응을 개시한다13,14.
TAS1Rs는
TAS1R1, TAS1R2, TAS1R3의 세 가지 멤버8,9로 구성되며,
이합체(heterodimer)를 형성하여
단맛(TAS1R2–TAS1R3 heterodimer) 또는 감칠맛(TAS1R1–TAS1R3 heterodimer)을 매개한다1,2,15,16 (Fig. 1a).
TAS1Rs 외에도 class C GPCR family에는
metabotropic glutamate receptors (mGluRs)17,
γ-aminobutyric acid receptors (GABABRs)18,
calcium-sensing receptor (CaSR)19,
그리고 여러 orphan receptors20가 포함된다.
TAS1Rs는 class C GPCRs의 특징을 공유하며, extracellular venus flytrap domain (VFTD), cysteine-rich domain (CRD), seven-transmembrane domain (TMD)으로 구성된다. 인간 단맛 수용체는 glucose와 sucrose 같은 천연 설탕, sucralose와 neotame 같은 인공감미료21, monellin, thaumatin, miraculin, brazzein 같은 천연 단백질성 단맛 물질22,23 등 다양한 단맛 자원을 인식한다. 주목할 점은 이 수용체가 설탕에 대해 micromolar에서 millimolar 범위의 낮은 결합 친화도를 보인다는 것이다24,25 (Fig. 1b). 단맛 인지는 에너지 대사와 밀접하게 연결되어 있는데, 이는 에너지-rich 음식으로 유도하는 초기이자 중요한 감각 신호 역할을 하기 때문이다26,27. 구강 미각 인지 외에도 TAS1R2와 TAS1R3 수용체는 장28, 췌장29, 뇌30에 ectopic하게 발현되어 대사 과정을 조절하는 데 관여한다.
따라서 ectopic TAS1Rs는 비만과 당뇨, 고지혈증 같은 대사 장애의 잠재적 치료 표적으로 부상하고 있다29,31,32. 화학감각 및 생리적 중요성에도 불구하고, 인간 단맛 수용체의 구조적 특성화는 여전히 큰 도전 과제이다. 이전 구조 연구에서는 medaka fish 유래 TAS1Rs의 VFTD crystal 구조만 보고되었으며33, 인간 TAS1Rs의 full-length 구조는 아직 알려지지 않아 감미료 조절과 수용체 활성화 메커니즘에 대한 이해를 제한하고 있다. 본 연구에서는 apo 상태와 sucralose-bound 상태의 full-length 인간 TAS1R2–TAS1R3 heterodimer에 대한 cryo-electron microscopy (cryo-EM) 구조를 제시하여, 분자 수준에서 단맛 인지를 이해하기 위한 구조적 템플릿을 제공한다.
Structure of apo- and sucralose-bound TAS1R2–TAS1R3 TAS1Rs pose challenges for structure determination owing to their low exogenous recombinant overexpression and the inherent instability of their heterodimeric complexes. To enhance expression, we utilized a fluorescent protein fusion strategy, fusing mScarlet3 protein34 and mNeonGreen35 with flexible linkers to the C terminus of TAS1R2 and TAS1R3, respectively (Extended Data Fig. 1a and Methods), and co-expressed the modified TAS1R2 and TAS1R3 using the BacMam system (Methods). Using these strategies, we obtained stable TAS1R2– TAS1R3 heterodimers in the apo and sucralose-bound states (Extended Data Figs. 1b,c, 2 and 3) and determined the cryo-EM structures at overall resolutions of 3.4 Å (apo) and 3.5 Å (sucralose-bound) (Fig. 1c,d and Extended Data Table 1). The EM density maps were of sufficient quality to distinguish TAS1R2 from TAS1R3, aided by the well-resolved sidechain densities in most parts of the VFTD (Extended Data Fig. 4a,c), unique glycosylation sites in the CRD of TAS1R2 (TAS1R2-CRD) (Extended Data Fig. 4b), as well as the TMD in the apo structure (Extended Data Fig. 4e,f). These electron microscopy maps enabled us to build structure models for most receptor regions. Notably, the electron microscopy density for sucralose in the binding pocket was well resolved. Additionally, we also determined the apo-TAS1R2–TAS1R3 structure in the absence of glycerol for the purpose of testing its potential activity on sweet taste receptor (Extended Data Fig. 5a–d and Methods), although our calcium mobilization assay showed that glycerol could not activate the sweet taste receptor even at 1.0 M concentration (Extended Data Fig. 1d). Consistently, this glycerol-free structure is identical to the apo structure (Extended Data Fig. 5e,f). Both heterodimer complex samples thus represent the true apo state of the receptor.
TAS1Rs는 외인성 재조합 과발현 수준이 낮고 heterodimeric complex의 본질적인 불안정성 때문에 구조 결정에 어려움을 겪는다. 발현을 향상시키기 위해 우리는 fluorescent protein fusion 전략을 사용하였으며, mScarlet3 protein34와 mNeonGreen35을 flexible linker를 통해 각각 TAS1R2와 TAS1R3의 C 말단에 융합하였다 (Extended Data Fig. 1a 및 Methods). BacMam 시스템을 이용해 수정된 TAS1R2와 TAS1R3를 공동 발현하였다 (Methods). 이러한 전략을 통해 apo 상태와 sucralose-bound 상태의 안정적인 TAS1R2–TAS1R3 heterodimer를 얻었으며 (Extended Data Figs. 1b,c, 2, 3), 전체 해상도 3.4 Å (apo)와 3.5 Å (sucralose-bound)로 cryo-EM 구조를 결정하였다 (Fig. 1c,d 및 Extended Data Table 1). EM density map은 VFTD 대부분에서 잘 해석되는 sidechain density (Extended Data Fig. 4a,c), TAS1R2-CRD의 독특한 glycosylation sites (Extended Data Fig. 4b), 그리고 apo 구조의 TMD (Extended Data Fig. 4e,f) 덕분에 TAS1R2와 TAS1R3를 구분할 수 있을 만큼 충분한 품질이었다. 이러한 전자현미경 map을 바탕으로 수용체 대부분 영역의 구조 모델을 구축할 수 있었다. 특히 binding pocket 내 sucralose의 전자현미경 density는 잘 해석되었다. 또한 glycerol의 sweet taste receptor에 대한 잠재적 활성을 테스트하기 위해 glycerol이 없는 apo-TAS1R2–TAS1R3 구조도 결정하였다 (Extended Data Fig. 5a–d 및 Methods). 그러나 calcium mobilization assay에서 glycerol은 1.0 M 농도에서도 sweet taste receptor를 활성화하지 못했다 (Extended Data Fig. 1d). 일관되게, glycerol-free 구조는 apo 구조와 동일하였다 (Extended Data Fig. 5e,f). 따라서 두 heterodimer complex 샘플 모두 수용체의 진정한 apo 상태를 나타낸다
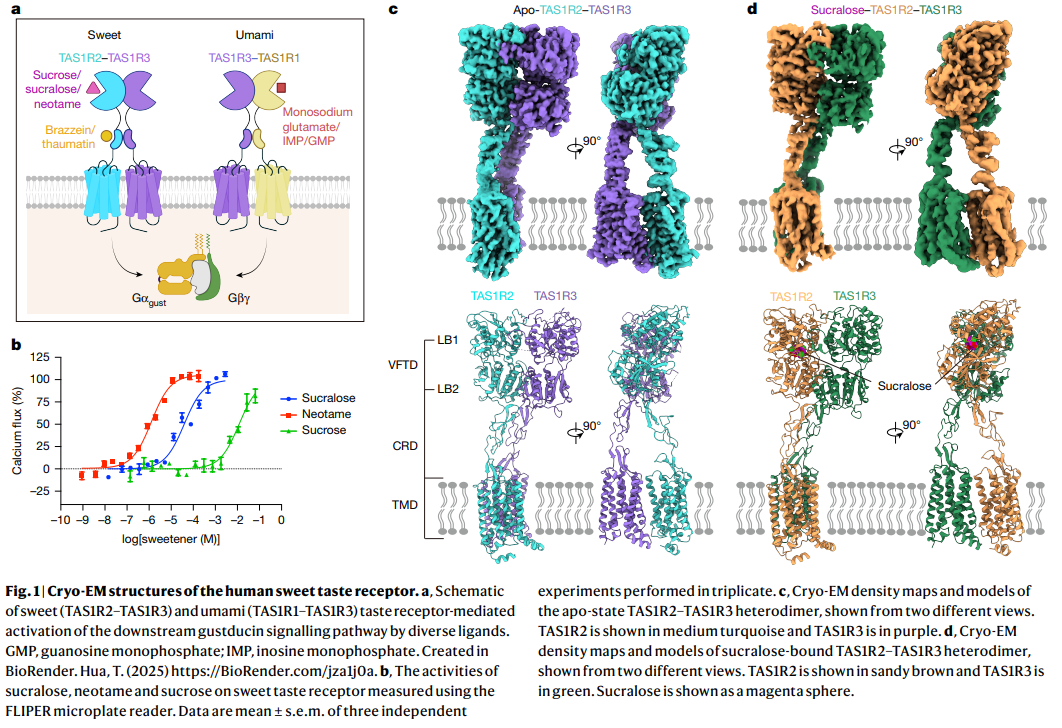
a. Schematic 개요
- 단맛 (Sweet): TAS1R2–TAS1R3 heterodimer. Sucrose, sucralose, neotame, brazzein, thaumatin 등의 다양한 단맛 물질을 인식.
- 감칠맛 (Umami): TAS1R1–TAS1R3 heterodimer. Monosodium glutamate, IMP/GMP 등.
- 두 heterodimer 모두 Gαgust와 Gβγ를 통해 downstream gustducin signalling pathway를 활성화.
b. Calcium flux assay
- Sucralose, Neotame, Sucrose의 농도에 따른 calcium flux (%). Sucralose와 Neotame이 sucrose보다 훨씬 강력한 agonist임을 보여줌.
c. Apo 상태 구조 (TAS1R2–TAS1R3)
- Apo (ligand-free) heterodimer의 cryo-EM density map과 모델.
- TAS1R2 (turquoise), TAS1R3 (purple)로 색상 구분.
- VFTD, CRD, TMD 영역 표시.
d. Sucralose-bound 상태 구조
- Sucralose가 결합된 heterodimer 구조.
- Sucralose (magenta sphere) 결합 부위 확인.
- TAS1R2 (sandy brown), TAS1R3 (green).
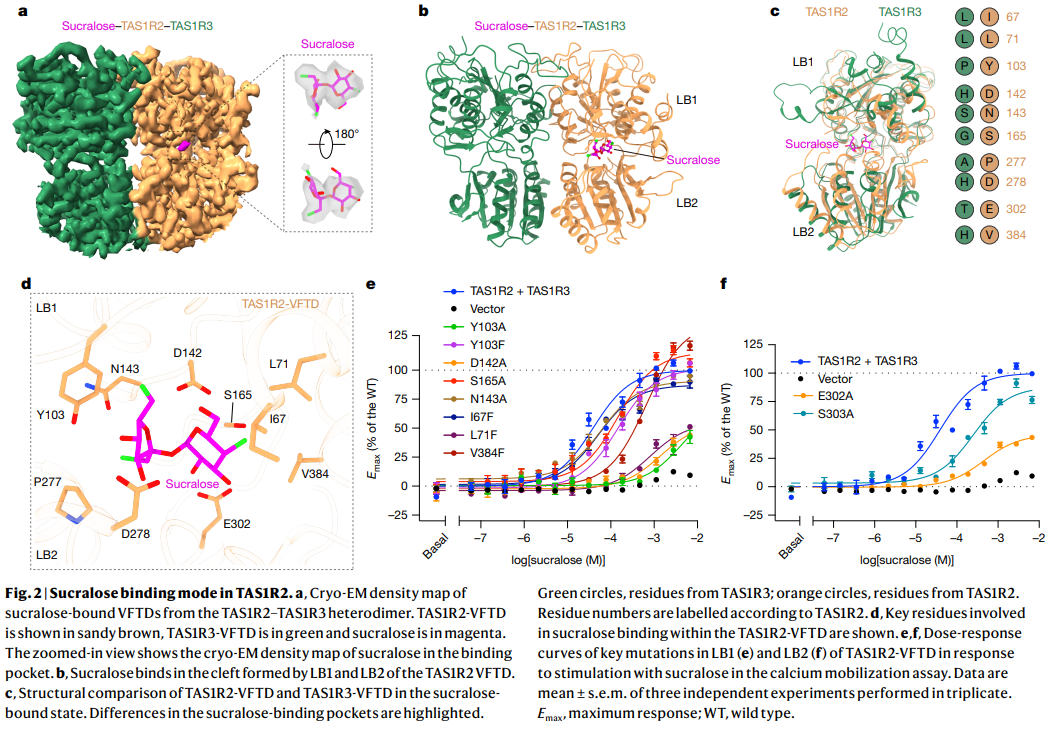
a. Sucralose-bound TAS1R2–TAS1R3 구조
- Cryo-EM density map에서 sucralose (magenta)가 TAS1R2-VFTD (sandy brown)에 결합한 모습.
- TAS1R3 (green).
b. Sucralose 결합 부위 확대
- Sucralose가 TAS1R2-VFTD의 LB1과 LB2 사이 cleft에 결합.
- 주요 결합 잔기 표시.
c. TAS1R2-VFTD 구조 비교
- LB1과 LB2 영역의 잔기 번호 표시 (TAS1R2: orange circle, TAS1R3: green circle).
- Sucralose 결합에 관여하는 핵심 잔기 확인.
d. Sucralose binding pocket 상세
- TAS1R2-VFTD의 LB1과 LB2 사이 cleft에 sucralose가 결합.
- 주요 상호작용 잔기: Y103, D142, N143, S165, I67, P277, D278, E302, V384 등.
e, f. 돌연변이 기능 분석 (Calcium mobilization assay)
- LB1 (e)과 LB2 (f) 영역의 핵심 잔기 돌연변이가 sucralose에 대한 dose-response curve에 미치는 영향.
- 여러 돌연변이 (Y103A, D142A, N143A 등)에서 최대 반응(E_max)이 감소하여 결합·활성화에 중요한 역할을 확인.
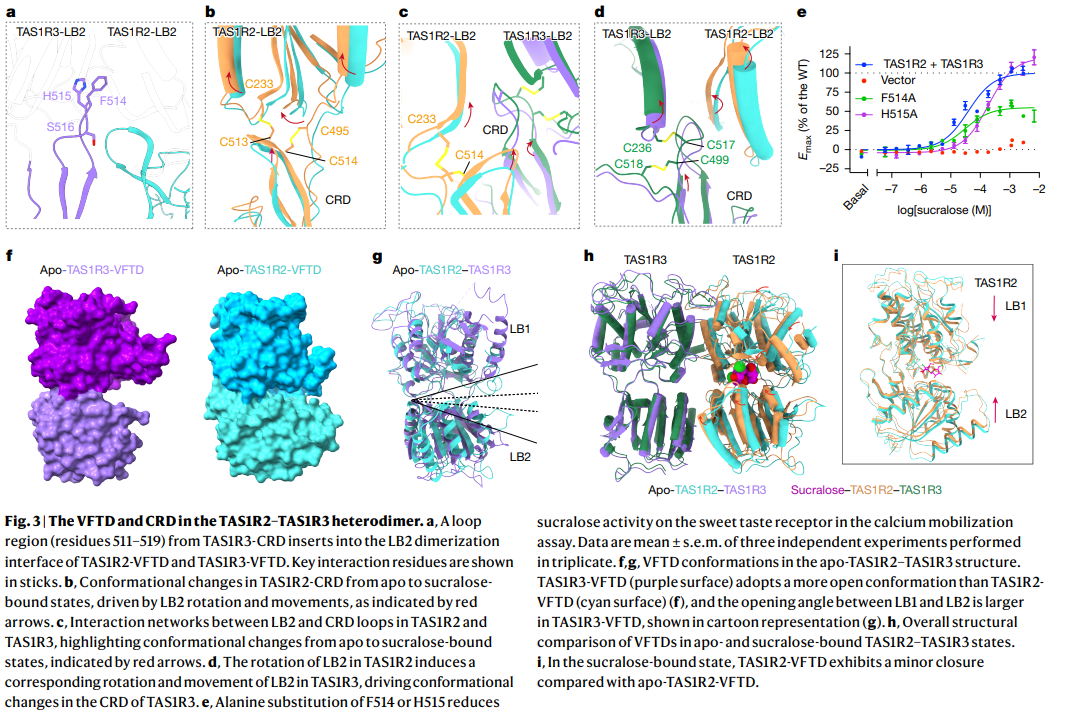
a. TAS1R3-CRD의 loop region (residues S11–S19)
- TAS1R3-CRD의 loop가 TAS1R2 LB2 dimerization interface에 삽입됨.
b. Apo vs Sucralose-bound 상태에서의 conformational change
- LB2 rotation과 movement에 의해 TAS1R2-VFTD와 TAS1R3-VFTD의 구조 변화 (red arrows).
c. LB2와 CRD loop 간 interaction network
- Sucralose 결합 시 TAS1R2와 TAS1R3의 LB2-CRD loop 간 상호작용 변화 (red arrows).
d. LB2 rotation in TAS1R3
- Sucralose 결합 시 TAS1R3 LB2의 rotation이 TAS1R2보다 크다.
e. F514A, H515A 돌연변이 효과
- Calcium mobilization assay에서 sucralose에 대한 반응 감소.
f, g. Apo 상태 VFTD 구조
- TAS1R3-VFTD (purple)와 TAS1R2-VFTD (cyan)의 표면 표현.
h. Apo vs Sucralose-bound 전체 구조 비교
- Sucralose 결합 시 전체 heterodimer의 구조 변화.
i. Sucralose-bound 상태에서의 TAS1R2-VFTD
- Apo 상태 대비 LB1–LB2 간 closing (minor closure) 관찰.
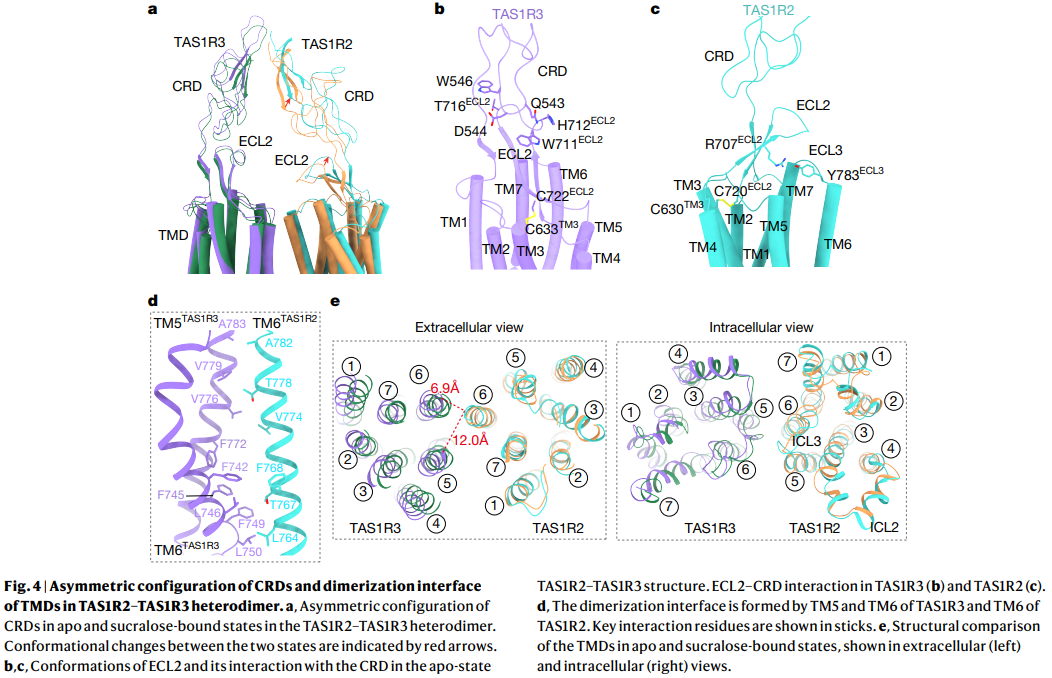
a. Apo vs Sucralose-bound 상태에서의 CRD 비대칭 구조
- TAS1R2와 TAS1R3의 CRD (cysteine-rich domain)가 비대칭(asymmetric)으로 배치됨.
- Sucralose 결합 시 CRD의 conformational change 관찰.
b, c. ECL2 (Extracellular Loop 2)와 CRD의 상호작용
- Apo 상태에서 TAS1R3의 ECL2가 CRD와 상호작용.
- TAS1R2와 TAS1R3의 ECL2 conformation 비교.
d. TMD dimerization interface
- TM5와 TM6 (TAS1R3)와 TM6 (TAS1R2)이 dimerization interface를 형성.
- 핵심 interaction 잔기 표시.
e. Extracellular vs Intracellular view
- Apo와 sucralose-bound 상태에서의 TMD 구조 비교.
- Extracellular view: helix 간 거리 변화 (6.9 Å vs 12.0 Å).
- Intracellular view: ICL (Intracellular Loop) 영역 구조 비교
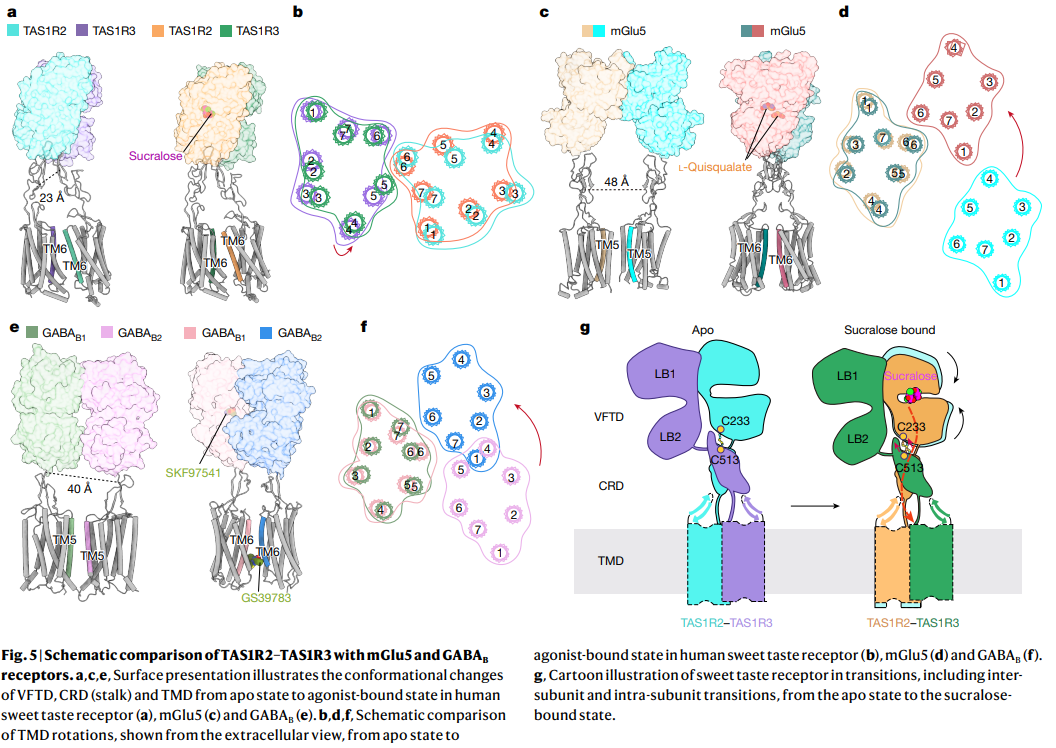
a–c. TAS1R2–TAS1R3 heterodimer의 구조 변화
- Apo 상태와 sucralose-bound 상태에서 VFTD, CRD, TMD의 conformational change.
- TAS1R2 (cyan)와 TAS1R3 (purple)의 TM6 rotation과 LB1–LB2 movement 관찰.
d–f. mGlu5와 GABAB 수용체와의 비교
- mGlu5 (agonist-bound)와 GABAB (GS39783-bound)에서도 유사한 TMD rotation과 dimer interface 변화 확인.
- TAS1R2–TAS1R3의 activation 메커니즘이 class C GPCR family의 일반적인 패턴임을 보여줌.
g. Cartoon 모델
- Apo 상태 → Sucralose-bound 상태로의 전이 과정.
- VFTD (LB1, LB2), CRD, TMD의 inter-subunit 및 intra-subunit 변화.
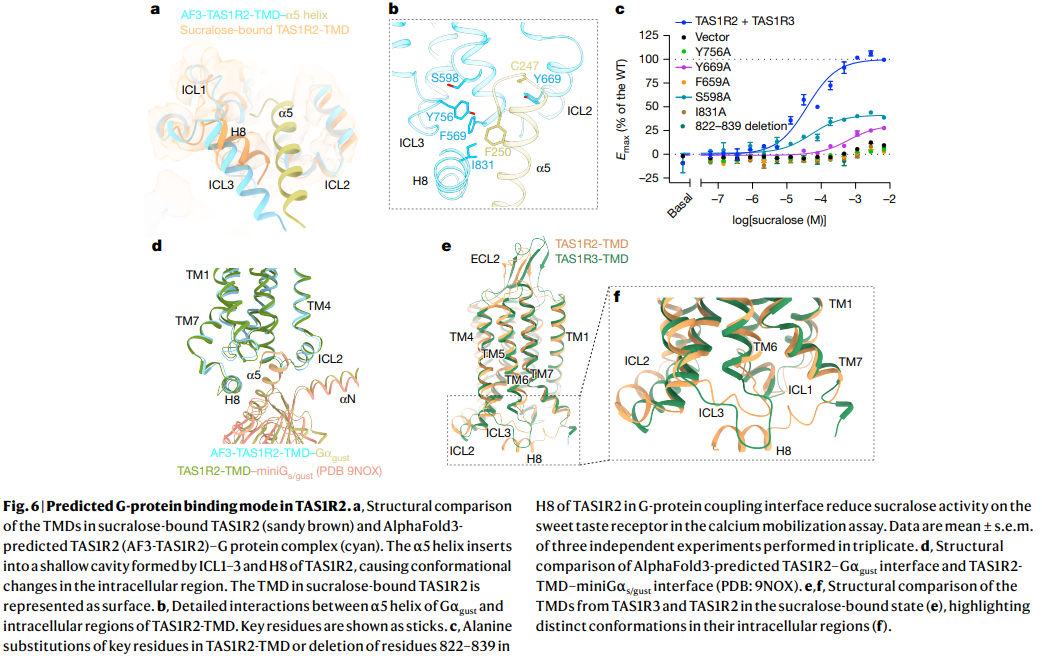
a. Sucralose-bound TAS1R2 TMD와 AlphaFold3 예측 G-protein 복합체 비교
- TAS1R2 TMD (sandy brown)와 AlphaFold3-predicted TAS1R2–Gαgust 복합체 (cyan)의 구조 비교.
- G-protein의 α5 helix가 TAS1R2의 ICL1–3과 H8 사이 cavity에 삽입되어 conformational change 유발.
b. TAS1R2 TMD–Gαgust interaction 상세
- ICL2, ICL3, H8 영역의 핵심 잔기 (S599, Y756, W669, F569, F250, I831 등)와의 상호작용 표시.
c. 돌연변이 기능 분석
- Calcium mobilization assay에서 TAS1R2 TMD 핵심 잔기 돌연변이 (Y756A, W669A, F569A, I831A 등)와 822–839 deletion이 sucralose에 대한 반응을 감소시킴.
d. TAS1R2–miniGαgust interface 구조
- AlphaFold3 예측 모델 (PDB: 9NOX) 기반 TAS1R2–G-protein coupling interface.
e, f. TAS1R2와 TAS1R3 TMD의 구조 비교
- Sucralose-bound 상태에서 TAS1R2와 TAS1R3의 TMD intracellular region의 distinct conformation 강조.
한 줄 요약: Sucralose 결합 시 TAS1R2 TMD가 G-protein (α5 helix)을 coupling하는 예측 모델과, 핵심 잔기 돌연변이가 receptor 활성에 미치는 영향을 보여주는 Figure