제11장 진화에 대하여 화석이 말하는 것
어떤 사람은 근본으로 가는 것을 완전히 잊는다. 어떤 사람은 중요한 문제를 문제로 삼지 않는다. - 루드비히 비트겐슈타인(Ludwig Wittgenstein)" -
화석은 '성경이나 과학이냐? 라는 오래된 문제에 대하여 우리에게 말해 줄 것이 많다. 화석은 진화라는 교리가 막다른 장벽을 만날 때, 애원할 수 있는 최후의 법정'으로 크게 환영을 받았다. 진화에 대하여 화석은 실제로 무엇이라고 말하고 있는가?" 화석이 진화를 지지한다는 주장은 정말로 옳은 것인가? 우리는 진화의 속도와 화석 집단의 연결이라는 두 가지 주요 문제에 대하여 살펴볼 것이다.
진화론적 변화의 속도와 화석 기록
수많은 종류의 복잡한 동물들이 대량으로 산출되는 지층의 바로 아래의 선캄브리아기 지층에서 나타나는 에디아카라 동물군과 버제스 셰일 동물군 등을 포함하여 대부분 선캄브리아기에 해당되는 매우 단순한 형태의 몇몇 중요한 화석들의 발견을 통해서, 시간의 흐름에 따라서 진화가 진행되었다는 일반적인 가설에 이의가 제기되고 있다. 이러한 가정도 기껏해야 진화 메커니즘의 속도는 매우 불규칙적이었다고 짐작하는 정도이지만 말이다.
진화 모델에 따르면, 생명은 최소한 35억 년 전에 출현하여 진화되었지만, 초기약 30억 년 동안은 단순한 단세포 상태로 있었다. 그 후 1억 년도 채 안 되는 사이에 갑자기 대부분의 동물계의 문(門, 약 40개)에 해당되는 동물들이 소위 '캄브리아기 폭발' 의 결과로 등장한다. 그 이후 새로운 동물문(門)이 진화(출현)한 것은 없다. 아무튼 캄브리아기 폭발에 1억 년이나 긴 시간을 할당하는 것은 진화론에 대한 관대한 취급이다. 어떤 이들은 대부분의 동물문이 발생하는 데 단지 5백만 년 내지 1천만 년을 제안하기도 하는데, 이것은 모든 진화 과정이 완전히 일어나는 데 요구되는 시간의 1/300보다 적은 기간이다. MIT의 바우링(Samuel Bowring)은 다음과 같이 비평한다.
"그리고 생물학자인 나의 친구들에게 잘 물어보는 말은, 생물이 자신의 불편함을 느끼기 이전에 진화는 얼마나 빠르게 일어나는가?"라는 질문이다." 또한 연구자들은 캄브리아기 지역에 조류가 두드러지게 많다고 보고하고 있다." 일반적으로 현생종(現)의 1/4에 해당되는 식물 화석이 상위 지층에서 다양한 생물군과 함께 발견된다<그림10.1>. 지질 주상도의 상부에서도 화석들은 계속해서 갑자기 출현된다. 예를 들어, 대부분의 포유동물 목(目)들은 지금으로부터 단 1,200만 년 전에 나타나는 것으로 보인다(제3기의 하부층). 진화론자 스탠리(Steven M. Stanley)는 평균적인 포유동물 종의 화석들은 연대적으로 100만년 이상 계속 나타나는데, 이 기간에 포유동물들이 고래나 박쥐처럼 상당히 변화된 모습의 또 다른 포유동물로 진화되기 위해서 10 내지 15차례의 계속적인 진화의 단계를 밟았을 정도의 시간밖에 되지 않는다고 지적하면서, 그는 "이는 너무나도 터무니없는 비상식적인 것"이라고 말하면서, "그렇게 짧은 기간에 매우 다양한 포유동물 형태가 갑작스럽게 출현된 것을 설명하기 위해서 조절 유전자의 신속한 변화의 가능성과 스스로 돌연변이를 가속화시킬 수 있는 작은 개체집단의 존재 가능성을 대안으로 제시하였다. 새들에 대해서도 5백만 년에서 1천만 년 사이에 현재 살아 있는 모든 조류목(鳥類目)들을 포함하는 '엄청난 폭발적인 진화가 일어났다고 계속 보고되어 왔다." 이미 우리는 단속평형설(punctuatedequilibria)이 해결책이 아니라고 지적하였다." 단속평형설은 분류학상의 종 수준의 변화를 다루는 것이지 목(目), 강綱), 문(門)과 같은 상위 분류 수준의 갑작스런 출현을 설명하는 것이 아니다.
실제로 화석 기록은 하부 현생이언 이후로 식물과 동물 모두의 기본 형태들이 점점 감소되고 있음을 암시한다. 굴드(Stephen J. Gould)는 현재보다 많은 동물의 기본형들이 캄브리아기 퇴적물에서 발견된다고 지적한다. 그는 현재의 해부학적 구도(anatomical plans)의 수가 과거보다 적기 때문에, 종래의 나무 둥치로부터 시작되어 점점 진화가 진행되면서 다양한 생물들(가지와 잎사귀)로 분화되는 전통적인 진화계통수進化, <그림 11.1>>가 거꾸로 그려져야 한다고 제안하였다. 10) 고식물학자 스튜어트(Wilson Stewart)와 로스웰(Gar Rothwell)은 현재의 23종류 식물군과 옛날 하부 고생대를 대표하는 31종류의 '주요 식물군'을 비교하여 목록을 작성하였다. 우리는 <그림 10.1>의 하부 고생대에서 훨씬 다양한 주요 생물종(生物들을 볼 수 있다. 고생대에서는 67종류의 식물군이 나타났으나 신생대에는 42종류만 보인다. 그림에서는 고생대의 더 작은 분류 단위의 식물군을 표시하지 않았기 때문에, 이러한 차이는 더 커질 수도 있다. 지질 주상도의 상부 지층에서 더 많은 종을 포함하고 있을지도 모른다. 2) 그러나 이것
은 기본형에 비해서 근소한 차이가 나는 변종들이 포함되기 때문이다. 다시 말해서 지질 주상도의 하부에서는 기본 주제곡이 많이 나타나고, 상부에서는 우세한 주제곡은 적으나 변주곡이 많은 것이다. 세월이 흐름에 따라 진화가 점차로 진행되면서 더 많은 종류의 출현을 기대하지만, 멸종 등에 의해서 지질 주상도의 상부에서는 본래보다도 더 적은 해부학적 구도(anatomical plans)만이 남아 있는 것이다.
불규칙적 진화 속도라는 말은 일단 변화가 일어나면 급속도로 진행되었을 것이라는 뜻이다. 화석은 캄브리아기 이전, 즉 전 지질 시대의 5/6 에 해당하는 기간에는 진화가 거의 없었다는 것을 나타낸다. 그 뒤에는 마치 신속한 진화의 변화와 변화 사이에서 빈번히 아무것도 없는 평형 상태가 진행되는 등 진화가 간헐적으로 일어나는 형식을 좇았을 것이다. 이대로라면 진화가 일어나는 과정에 소요된 실제적인 시간은 아주 짧았다는 결론이 나온다. 어떤 진화 모델에 따르면 진화에 사용된 시간은 지질 시대의 단 1% 기간에 불과하다. 만일 화석 기록에서 이러한 패턴을 정말 나타내고 있다면 전체 진화 과정에 필요되었을 것으로 제안하고 있는 수십억 년의 시간을 현저하게 감소시키는 결과를 가져올 것이다. 과거에는 진화에 필요되는 시간의 부족 때문에 진화론에서 당면하는 많은 불가능성을 한층 고조시켰기 때문이다. 13)
14)그러나 진화론자들은 화석 기록을 통해서 급격한 변화를 기대하지만, 실제로 얻은 자료는 진화 발달이 극히 느린 속도로 진행되었다는 심증만 확인할 뿐이다. 현재 살아 있는 일부 생물들은 화석과 모양이 거의 일치한다. 4억 년 전에 진화를 완료한 것으로 가정되는 하부 데본기의 진드기는 현생종과 거의 동일하다. 쇼프(J. William Schopf)는 호주 중앙부의 비터스프링스 암석층으로부터 추정연대 8억 5천만 년 전의 남조류(남세균)로 추정되는 여러 화석 표본을 발견하였는데, 현재 살아 있는 종들과 너무나 똑같다. 그는 또 추정연대는 각각 다르지만 현생종과 똑같은 90여 종의 화석에 대하여 보고하였다. 13) 후기 시생대에서 중기 원생대(지금부터 12~27억 년 사이)에서 이와 비슷한 생명체에 대하여 의견을 피력한 스튜어트(Wilson Stewart)와 로스웰(Gar Rothwell)은 "비록 이것들의 생물학적 체계의 진화 속도에 대해서는 아무것도 결정된 바가 없다 하더라도, 선캄브리아기로부터 현재까지 이들의 형태학적인 모습은 하나도 변치 않고 겉보기에 완전히 일치하고 있다."고 말한다.“
북미 오대호 지역의 20억 년 전의 지층으로 가정하는 건플린트 쳐트층(급)에서 발견되는 형태들도 역시 현재 살아 있는 생물종과 거의 동일하다. 좀 더 일반적으로 말하면, 놀(Andrew Knoll)은 "많은 후기 원생대의 원핵생물(原核生物,
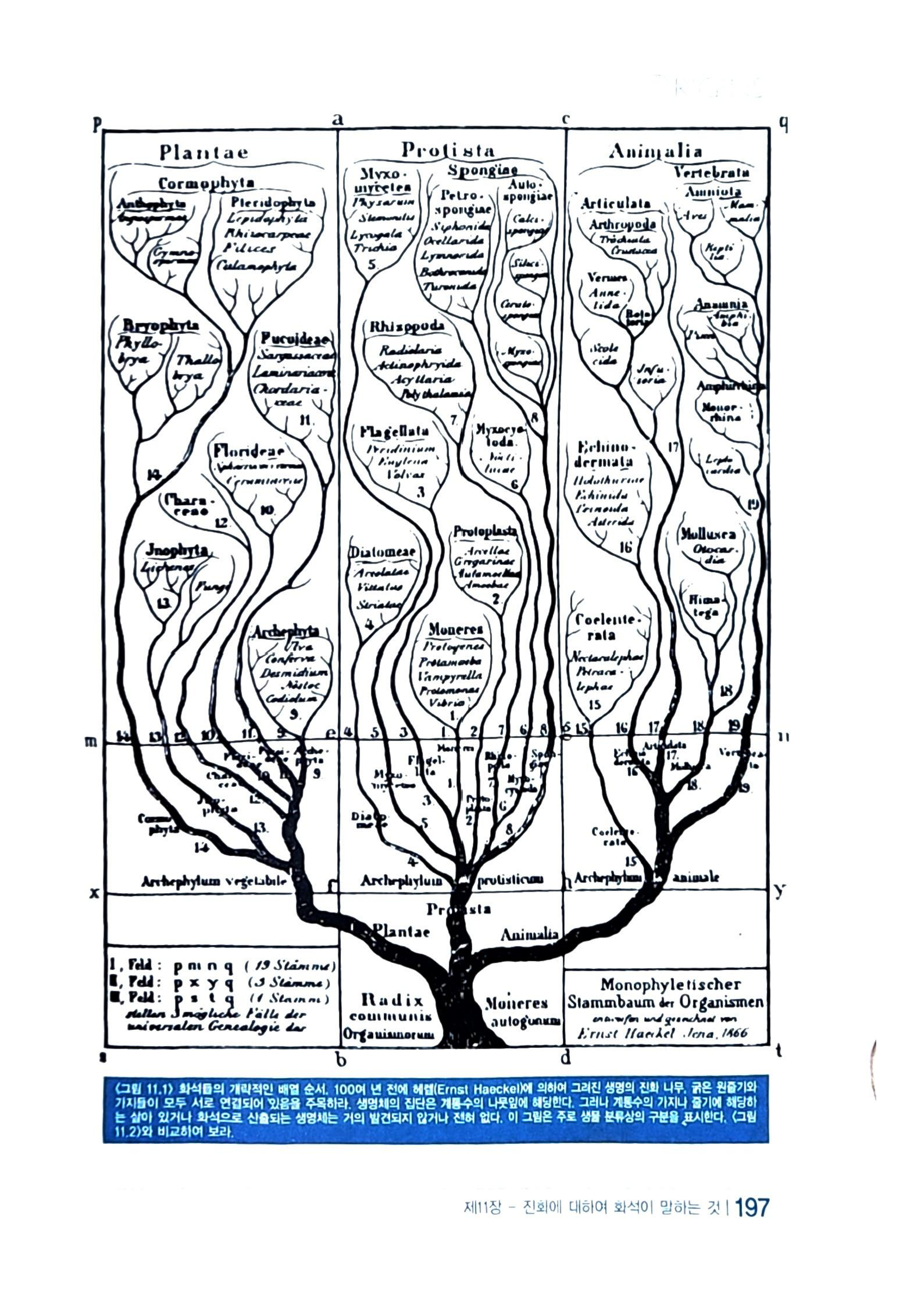
<그림 11.1> 화석들의 개략적인 배열 순서, 100여 년 전에 헤렐(Ernst Haeckel)에 의하여 그려진 생명의 진화 나무, 굵은 원줄기와 가지들이 모두 서로 연결되어 있음을 주목하라. 생명체의 집단은 계통수의 나뭇잎에 해당한다. 그러나 계통수의 가지나 줄기에 해당하는 살아 있거나 화석으로 산출되는 생명체는 거의 발견되지 않거나 전혀 없다. 이 그림은 주로 생물 분류상의 구분을 표시한다. <그림 11.2)와 비교하여 보라.
원시적인 세포핵을 가진 생물)은 현재 살아 있는 남조세균과 형태, 발생, 또는 습성과 다른 것이 거의 없다."고 말한다. 진화론자는 이러한 불변(不變)의 원인을 진화의 우연적(불규칙적) 속도, 또는 눈에 보이지 않는 내부적 변화에 근거하여 설명하려고 노력한다. 그러나 창조론적 맥락에서 보면 이러한 유사성은 최근의 살아 있는 생명체가 암석 속으로 침투한 결과일지도 모른다고 보고 있다. 18)
진화를 화석 기록에 맞추기 위해서 진화가 극히 느리게, 또는 아주 빠르게 진행되었다는 등의 가정을 하는 것은 진화의 일반적인 이론이 현재 수시로 변하고 있는 자료에 어떻게 잘 대응해 가고 있는지를 보여 주고 있다. 진화가 매우 빠르게 진행되었다는 빠른 진화 속도는 느리고 점진적인 진화라는 전통적인 관념에 많은 도전이 되고 있다. 그리고 사람들은 왜 일부 세균이나 이와 비슷한 단순한 생명체가 6억 년 내내 진화하여 인간으로까지 진화한 반면에, 왜 일부 생명체들에게는 과거 20억 년간 전혀 변화가 나타나지 않았는지 이상하게 생각한다. 진화 모델에 대해서 화석은 기껏해야 고도의 불규칙한 진화 속도를 나타내준다. 화석에서 보여 주는 오랜 기간의 느린 진화 혹은 진화가 없어 보이기까지 하는 긴 기간은 전혀 가능성이 없어 보이는 복잡한 진화에 조금도 시간을 할애해주지 않는다.
화석 기록에서의 시간 공백
필자가 대학원 학생이었을 때, 진화론 담당 교수가 필자의 창조론 신념에 대하여 동물학과 교수들이 염려하고 있다는 말을 전했다. 그 교수는 내가 창조론에 대해서 어떻게 설명하는지 궁금하게 생각하였다. 그래서 나는 교수에게 어떻게 하면 내가 창조론에서 진화론으로 돌아설 수 있는가를 보여 주었고, 내가 갖고 있는 몇 가지 진화론에 대한 의문점들을 질문하였다. 교수는 많은 관심을 갖고 나에게 대하였다. 필자가 제시한 요점 중 하나는 거북이는 어떻게 중간 형태의 화석을 남기지 않고 파충류로부터 진화할 수 있는가' 하는 것이었다. 거북이는 독특한 생명체이다. 이같이 특이한 거북이의 등판이 조개처럼 단단하게 진화될 때 많은 중간 형태를 거쳐서 진화되었을 텐데 화석 기록에는 그러한 증거가 전혀 없다. 고생물학자들은 크기 약 4m짜리를 포함해서 수천 마리의 거북 화석을 발견하였다. 진화론자들은 거북이가 2억 년 이전부터 진화되었다고 추정하지만, 거북이 화석이 처음으로 산출되는 지층 아래에서는 거북이가 점진적으로 진화되었다는 진화계통을 우리는 하나도 찾아볼 수 없다. 1) 그 외의 다른 주제들에 대해서도 토론한 후, 교수는 나의 대답에 만족한 듯이 보였고 진화론에는 여러 문제가 있다고 동의
하였다. 후에 들으니 필자가 학위를 받도록 허락된 단 하나의 이유는 나에 대한 교수들의 의견이 통일되지 않았기 때문이라고 했다.
거북의 기원과 같은 문제는 수백 번 반복될 수도 있다. 지질 주상도의 연속되는 각 층에서 우리는 새로운 종류의 생명체가 갑작스럽게 출현하는 것을 많이 본다. 하부 지층에서 그것들의 조상을 찾으려는 노력은 아직 성공하지 못하였다. 다윈(Charles Darwin)은 이러한 문제를 충분히 알고 있었다. <종의 기원>에서 다윈은 "그러나 멸종이 거대한 규모로 작용되었다면 이러한 비율로 그 이전의 중간 형태의 변종도 참으로 거대했을 것이다. 그렇다면 왜 지질층에는 그러한 중간 형태의 연결 종들로 가득 채워져 있지 않는가? 지질학은 확실히 그렇게 정교하고 점진적인 유기체의 사슬을 나타내지는 않는다. 그리고 이것은 아마 (자연선택) 이론에 대하여 주장될 수 있는 가장 분명하고 중대한 결함이다."라고 말했다.201 다윈은 이 문제의 원인을 지질 기록의 '극도의 불완전성' 때문이라고 탓하였다. 그러나 그 당시에 최고의 화석 권위자로 다윈 자신도 인정한 아가시즈(Agassiz), 픽테트(Pictet), 세지윅(Sedgwick) 등은 다윈의 견해에 반대하였다. 21
중간 형태의 누락 사실은 다윈 시대나 지금이나 뚜렷하게 변하지 않았다. 120년 후 미국 시카고 필드 자연사 박물관(Field Museum of Natural History)의 지질학 분야 관장이며 미국 고생물학회의 회장이었던 라프(David M. Raup)는 다음과 같은 사실을 관찰하였다. 다윈 시대나 오늘날의 지질학자들이 실제로 찾아낸 것은 생명체의 점진적인 진화가 아니라 대단히 불균일하거나 변덕스런 기록일 뿐이다. 즉 종은 매우 갑자기 출현해서, 자신들이 존재하는 동안 조금도 변화를 보이지 않다가, 그 다음에는 기록으로부터 돌연히 사라진다."22)
그리고 불과 수년 전에 오크라하며 대학교의 고생물학자 키츠(David B. Kitts)도 이를 인정하였다. “고생물학이 눈에 보이는' 진화의 수단을 제공해 줄 것이라는 빛나는 기대에도 불구하고, 고생물학은 진화론자들에게 불쾌한 문제들을 안겨줬는데, 그것은 화석 기록에서 '시간 간격(gaps)'의 존재는 진화론자에게 악명 높은 치명적인 문제가 되고 있다. 진화는 종(種)들 사이의 중간 형태를 요구하지만 고생물학은 중간 형태를 하나도 제공하지 못한다."23)
굴드(Stephen Jay Gould)는 되풀이하여 "화석 기록상에서 전이형태 )가 아주 희귀한 것은 마치 고생물학에서 어떤 '업계의 비밀' 처럼 지속되어 왔다. 교과서에 나오는 진화 계통수를 증명해 줄 수 있는 자료는 마디가 갈라지는 곳과 잔가지에 양념을 치듯 극소량의 자료뿐이고, 나머지는 화석에 근거된 자료가 아니다."라고 상기시켰다. 21)
화석의 배열 순서의 패턴들은 진화가 돌발적이고 급속도로 발생되었다고 말하
도록 진화론자들을 부추겼다. 진화론자들은 또한 진화가 중간형태의 화석으로 보존될 기회가 적은 작은 개체군 집단에서 발생하였을 것이라고 가정한다. 즉, 단속평형설을 신봉한다. 25) 이러한 설명은 가까운 동족(同族)의 종 사이에 있는 중간형태가 발견되지 않는 원인에 대해서는 잘 설명될지 모르지만, 주요 생명체의 개체군 사이에 있는 커다란 중간형태의 누락이라는 훨씬 더 중요한 문제를 설명하지 못한다.
살아 있는 생명체와 화석 생물체는 문(門, phyla, divisions)이라고 부르는 주요 분류 계급으로 나눈다. 이것들은 계통적 분류 체계에 있어서 분명히 주요한 분류 단위이다. 80개 정도의 문(門)으로부터 갈라져서 현재 백만 종이 넘는 살아 있는 현생종(種)들을 형성하고 있다. 왜 큰 분류 단위인 문(門)이 이처럼 확실하게 구분되어져야 하는가? 그리고 우리가 화석을 고찰할 때, 왜 주요한 생물군들 사이에서 진화된 중간형태를 발견하지 못하는가? 이러한 확인 과정에서 진화 모델은 가장 치명적이고 결정적인 결함을 갖게 된다. 하나의 기본 종에서 다른 기본종으로의 변형과 같은 일종의 진화의 기적에 대한 희망이 아무런 실례도 없이 그냥 남아 있다.
고생물학자들은 앞으로도 새로운 화석종(化石들을 발견할 수 있겠지만, 과거수 세기 동안의 경험처럼 이것들은 중간형태가 아닌 새로운 문(門)으로, 또 하나의 새로운 분류군에 속하는 생물로 분류될 것이다. 26) 사람들은 다윈이 그랬던 것처럼, 화석 기록이 불완전하지만 수백만 개의 화석들이 수집되었다고 주장할지 모른다. 이 화석들이야말로 분류학상으로 문(門)과 같은 개체군으로 계속 분류되어야 할 화석들이고, 또 이 화석 속에는 진화론자들이 아직도 설명할 수 없는 큰 간격)이 그대로 남아 있는 것이다. 주요 개체군 사이에 진화가 발생하지 않았을 때만 화석이 형성되고 보존되기에 유리한 격변이나 돌발적인 사건이 발생했을 것이라고는 보지 않는다.
하버드 대학교의 고생물학자 심프슨(George Gaylord Simpson)은 분류 체계의 상위 단계로 올라가면 중간단계의 수가 감소하는 데 대한 문제점을 기술하였다. 그가 평가한 내용을 <표 11.1>에 나타내었다. 2 진화 모델에 따르면, 우리는 중간단계가 없는 주요한 생물 개체군들 사이에서 수많은 중간단계를 볼 수 있어야 한다. 잃어버린 연결고리(missing links)의 문제를 몇 가지 예를 들어 설명하려 한다. 28) 캄브리아기 폭발은 모든 주요 동물문(門)이 지질 주상도상의 같은 곳에서 등장하는 것은 아니다. 캄브리아기 폭발은 생물이 어떻게 어디로부터 진화되었는가를 보여줄 만한 선조나 조상(祖上) 생물이 없는 상황이다. 고생물학자들은 캄브리아기 폭발 바로 아래의 지층, 즉 중간형태를 발견할 것으로 기대되는 암석층을
철저히 연구하였다. 이러한 조사는 사실상 쓸데없는 일이었다. 화석으로부터의 증거가 부족한 가운데, 고생물학자들은 생물 개체군들 사이의 유연관계를 규명하는 일에 큰 곤경을 느낀다. 스크립스 해양연구소의 쉬람(Frederick Schram)은 "지금까지의 연구 제목 중에서 무척추동물 문(門)들의 연관성을 다루는 주관적인 이론이 관심을 끈 경우는 없었다. 이 분야에서 전문연구원 2명이 있다면 이들은 서로의 의견이 일치되지 않는다. 게다가 무척추동물 해부학에 대한 개인적인 관점에서 본 수많은 해석과 갖가지 방법으로 가상적(假的)인 조상이나 혹은 이론상으로만 있는 동물에 적용시킨 혼란스러운 명칭들은 가히 아찔할 정도이다."라고 말한다. 29)
식물의 진화에 대한 것도 크게 다르지 않다<그림 10.1>, 텍사스 대학교의 볼드(Harold C. Bold) 등은 다음과 같이 언급하였다. "현재 이용할 수 있는 비교형태학, 세포학, 생화학 그리고 화석 기록의 증거들을 주의 깊게 비교 분석한 후에, 그들은 식물계를 임시로 19개의 문(門)으로 분류했는데, 이 중에서 현재 2~3개의 문(門)을 통합하려고 하지도 않는다.""30)
예를 들면 화석 기록에서 현화식물(顯植物, 꽃피는 식물들은 갑자기, 완전한 형태로, 무수히 나타난다. 다윈은 현화식물의 기원을 '지긋지긋한 불가사의'라고 불렀다. 그로부터 한 세기가 지난 후에도 고생물학 선두주자들(Axelrod, Bold, Knoll, and Rothwell)은 역시 이것을 여전히 지긋지긋한 불가사의' 라고 불렀다." 날아다니는 생물은 크게 나누어 곤충, 익룡(날아다니는 파충류), 새, 박쥐 등 네 종류가 있다. 날아다니려면 날개 외에도 많은 기능이 필요하다. 예를 들어 경비행기의 구조는 자동차와는 명백히 다르다. 우리는 화석 기록상에 남아 있는 점진적인 진화의 증거인 날개를 당연히 기대할 것이다. 그러나 지질 주상도에 처음 나타날 때부터 화석 곤충은 비행 기능이 완벽하게 발달된 상태였다. 32) 또한 나는 익룡, 새, 박쥐도 완벽하게 비행 (飛行)이 가능한 생명체로 느닷없이 나타난다.

<표 11.1> 분류표에
생물체가 날아오르는 데 필요한 해부학상의 변화에는 골격, 근육조직, 깃털, 호흡, 신경계통의 형질 전환 등이 포함된다. 이러한 변화는 오랜 시간을 필요로 할 것이다. 그리고 이 같은 변화를 받은 생명체는 분명히 중간형태를 남길 것이다. 진화론에서 새의 깃털은 선조형(先祖) 파충류의 비늘에서 진화했을 것으로 가정한다. 현미경으로 깃털을 자세히 조사한 사람은 대단히 복잡하고 고도로 분화된 구조임을 깨닫는다. 진화를 지시하는 자도 없고 또 정해진 방향도 없는 진화 과정에서 때로는 실패하는 진화 계열도 있는데, 이러한 모든 것을 포함해서 파충류의 비늘로부터 날개의 모든 세부 부분에 이르기까지 이러한 모든 부분을 생기게 하는 집중적인 진화 과정이 암석 속의 화석 기록으로 왜 남겨지지 않았을까? 아직까지도 알려진 것은 하나도 없다.
잃어버린 연결고리
화석 기록이 기본적으로 불연속적이라는 사실에도 불구하고 몇몇 생명체는 잃어버린 연결고리를 나타내는 것처럼 보인다. 이런 생명체들은 진화 계열의 간격을 메우는 중간단계로 간주된다. 그래서 당연히 진화론자들은 일반인들이 그러한 예를 무시하지 않는다는 것을 확인하고 싶어 한다. 가장 유명한 예는 대부분의 생물학, 고생물학 책에 실린 파충류-새'인 시조새 (Archaeopteryx)이다. 다윈이<종의 기원>을 출판한 지 2년 후 독일에서 발견된 시조새는 해부학적으로도 중간 형태이고 지질 주상도에서도 바로 그곳에 위치하여 진화라는 개념을 확고히 하는데 도움을 주었다. 시조새는 파충류의 특징인 이빨, 긴 꼬리, 날개에 달린 발톱들 그리고 약간의 파충류를 닮은 골격 형태를 지니고 있다. 또 이는 조류의 특징인 완전히 발달된 깃털, 새의 가슴뼈 그리고 움켜쥐는 발톱 등을 가지고 있다. 33 시조새에서 발견되는 파충류의 어떤 특징은 파충류에만 있는 것이 아닌 것도 있다. 화석으로 나타나는 많은 새들이 이빨이 있고, 또 살아 있는 새들 중에는 날개에 발톱이 있는 종류도 있다. 시조새에서 볼 수 있는 완전하게 발달된 날개 깃털은 시조새가 틀림없이 조류임을 확인해 준다. 3) 시조새는 아마 파충류의 특징을 지닌 새였을 것이다. 최근 조상형 '새'로 발견된 것이 2종류가 있긴 하다. 그러나 깃털이 전혀 나 있지 않다. 한 마리는 지질 주상도상으로 보아 시조새와 거의 같은 지층에서 발견되고, 다른 한 마리는 조금 아래의 지층에서 발견되었다. 이들에 대해서는 아직 심한 논란 중에 있다. 35)
교과서에는 점진적인 진화를 예증하는 말 화석 시리즈' 그림이 가끔 나온다. 창조론자들은 진화론자들이 변화(진화)라고 주장하는 내용들이 너무나도 사소한
것들이고 또 창조된 생물들이 큰 종류들 사이의 중간형태에 해당하는 변화라고 볼 수 없기 때문에 그러한 주장에 그다지 관심을 기울이지 않는다. 그럼에도 근래에 진화론자들은 마르쉐(O. C. Marsh)에 의해 작성된 말 화석 진화도' 의 타당성에 대하여 문제를 제기하고 있는 것을 보면 큰 흥미를 느끼게 된다. 36) 심프슨은 다음과 같이 말하였다. "'측면 발가락 수의 점진적인 감소'라는 유명한 말의(정향)진화 경향은 단지 허구일 뿐이다.""37)
라우프(Raup)는 더 나아가 다음과 같이 진술한다. "진화의 기록들은 놀랍게도 여전히 단속적이다. 아이러니하게도 지금은 다윈 당시보다도 진화론적인 변이의 증거가 훨씬 더 줄어들었다. 여기서 내가 뜻하는 바는 북미에서의 말의 진화와 같은 과거의 다윈의 변이를 말해 주는 보기들 중 얼마는 파기되거나 아니면 보다 상세한 새로운 자료로 수정되어야 한다는 것이다. 왜냐하면 상대적으로 유용한 자료가 거의 없었던 때의 단순 명료한 발달(진화)의 좋은 본보기로 비쳐졌던 것들이 이제는 더욱 복잡하면서도 덜 점진적인 것으로 여겨지고 있기 때문이다.'"38)
미국 자연사 박물관에 전시되어 있던 말의 진화'의 원본은 이제 전시장에서 철거되었다. 30) 학자들은 말의 진화에 대한 새로운 개념을 구상 중이며, 최근의 견해로는 좀 더 연구가 필요하다는 것이다.1)
진화론자들은 가끔 단궁류(synapsid)라고 부르는 포유동물과 파충류 사이의 멸종된 중간형태에 대하여 언급한다. 진화론자들은 한 개체군의 다양한 골격형태를 다른 개체군과 연관이 있는 것처럼 관련시켰고, 단궁류 턱의 몇 특징들을 제한적이기는 하지만 포유동물과 파충류 사이의 진화계열의 좋은 본보기로 삼았다. 옥스퍼드의 고생물학자 켐프(T.S.Kemp)는 이것을 가리켜 "참으로...동물계에서 실제 화석 기록에 의하여 약간의 증거가 보존되어 있는 유일한 중간형태"라고 진술하였다. 그러나 개체군은 매우 다양하다. 단궁류의 어떤 특징은 그들의 선조(先祖)가 포유류라고 하는 기준에는 잘 맞지만, 다른 특징들은 이 기준에 맞지 않는다. 비록 일부의 특징들이 중간형태에 해당되기는 하지만, 포유류와 파충류 사이의 후손이라는 확신을 주지 못한다. 캐나다 맥길 대학교의 고생물학자 캐롤(Robert Carroll)은 "우리는 아직 포유동물에 이르는 구체적인 계보를 밝혀낼 수 없다."고 하였다. 12)41)
진화론자들은 잃어버린 연결고리의 또 다른 예를 제시한다. 어떤 사람들은 고래의 기원에 관한 계열을 제시한다. 일반적으로 주요 개체군 사이의 간격(gaps)을 연결하는 데 필요한 잃어버린 연결고리는 수십만 개로 추정되는 데 비하여 제시된 잃어버린 고리의 수는 극도로 적다. 아주 적은 보기를 강조하는 것은 증거 자
데본기
<그림 11.2> 진화 나무의 한 부분으로 양서류 화석에 기초한 최근의 도표 가정한 중간형태의 화석이 거의 나타나지 않음을 주목하라. 이는 화석 기록에서 중간형태가 없다는 사실을 반영한다. 오히려 과(科)' 수준으로 볼 때 신생대보다는 고생대에서 양서류가 더 많다. 해된 에리오포이데아 집단은 12개 과를 포함한다. 신생대 집단에서 하나인 것과 비교하라(Carroll, p.157 (참고문헌 190 참고, W.H.Freeman and Co. 출판사의 허가를 얻어 사용하였다.).
료가 그만큼 없다는 것을 드러내는 것이다. 심지어 진화의 연결고리라고 주장되는 화석을 보면 가장 큰 간격인 문과 문 사이의 수준에는 조금도 미치지 못하는
것들이다. 이미 언급했듯이 고생물학자들은 지금까지 대략 80개 정도의 문(門)과 이보다 더 많은 아문(門)으로 생물들을 분류하였고, 25만 종 이상의 화석들을 확인하였다. 그런데 이 중에서 단지 몇 점의 화석만이 중간형태로 간주되는 것을 보면 이것이야말로 진화는 심각한 문제점을 갖고 있음을 알 수 있다.
앞에서 우리는 생물종들이 단순한 생명체에서 복잡한 생명체로 진화된 것이라고 상상하고, 진화의 경로를 묘사하기 위해서 진화론자들이 제작한 진화 계통수에 대해서 언급하였다. 그러나 화석 집단들 사이에 어디서든지 존재하는 간격(gaps) 때문에 여러 가지 상이(相)한 배열이 가능하다. 진화 계통수에 관한 어떠한 포괄적인 안(案)도 서로 일치되는 경우가 드물다. 오히려 그러한 계통수를 보면 줄기나 가지에 해당하는 실제 생명체가 없기로 소문이 나 있다. 그리하여 최근에 고생물학자들은 더 조심스러워하며, 진화 계통수의 미확인된 부분을 이제는 불확실한 것으로 확인하고 있다.
<그림 11.1>은 유럽에서 진화를 열렬히 옹호했던 헤켈(Ernest Haeckel)의 선구자적 업적에 기초한 1886년도 진화 계통수이다. 그림에서 모든 것이 완벽하게 잘 연결되어 있는 것을 보게 된다. <그림 11.2>는 1888년에 작성된 양서류의 화석 기록에 관한 도표이다. 대부분의 개체군 집단이 서로 연결되지 않았음을 주목하기 바란다. 화석집단의 불연속성은 진화가 아닌 창조를 지지한다. 우리는 진화에서는 주요 집단들이 모두가 서로 연결되어 있을 것으로 기대한다. <그림 10.1>은 생물학자들이 생물들을 분류해 놓은 주요 개체군(집단)들을 보여 준다. 만일 진화가 실제로 발생했다면 각 집단은 화석 기록의 하부층에서 발견된 중간형태들을 사이에 두고 서로 연결되어야만 한다. 그러나 다시 말하지만 여기에는 중간형태가 하나도 없다.
화석 기록의 완전성
때때로 진화론자들은 중간형태의 생명체들은 몸이 부드러운 물질로 이루어져 있어서 단단한 물질로 이루어진 생명체처럼 쉽게 보존될 수 없기 때문에 중간형태 화석들이 날아가 버린 것이라고 주장한다. 43) 하지만 부드러운 물질을 지닌 생명체도 화석으로 잘 보존되어 있기 때문에 이러한 주장은 별로 의미가 없다. 진화론에 있어 가장 커다란 난제 중 하나는 캄브리아기 폭발이다. 캄브리아기 하부의 에디아카라 동물군과 캄브리아기의 버제스 셰일 동물군은 주로 연질의 몸체이지만 많은 수가 아주 잘 보존되어 있다. 반면에 캄브리아기 폭발 이전의 것으로 발견이 예상되는 중간형태의 생명체는 아직도 잃어버린 상태이다.
44) 또한 진화론자들은 화석화(化石化) 작용의 조건을 갖추지 못하여 화석 기록이 불완전하다고 주장한다. 그러나 화석 기록은 이전에 생각하던 것보다 훨씬 완벽하다. 개개의 생명체가 화석이 될 가능성은 희박하지만, 어떤 종의 총 개체수는 충분히 많아서 진화 기간에 걸쳐 종들을 대표할 화석이 잘 보존되는 일은 얼마든지 있을 수 있는 일이다. 동일 지역에서 화석으로 보존된 종류와 현재 살고 있는 종류를 비교하는 최근의 다양한 연구에 의하면 (개개의 생물체가 아닌) 종의 보존비율은 매우 높게 나타난다. 이와는 관련이 적은 다른 연구 결과에서도 같은 결론이 나왔다. 일반적으로 연체동물이 화석으로 나타나는 확률은 83~95%로 추정된다. 대합조개나 달팽이의 경우 77~85%이고) 개형충류(껍질이 있는 갑각류)는 60%이다. 많은 작은 집단으로 구성되는 보다 상위 분류 체제의 집단에서는 당연히 더 잘 보존된다. 육상 척추동물의 목(目) 수준에서는 98%까지 화석 기록이 보존되며, 과(科) 수준에서는 79%까지 보존된다." 이러한 양상은 화석 기록이 상당히 완전하고, 다윈의 주장처럼 그렇게 불완전하지 않다는 것을 보여 준다. 이것은 화석 형태들 사이에서 보이는 간격은 본래부터 존재하는 간격이라는 것을 암시하고 있다. 46)
사람들 마음에 깔려 있는 의식(意識)
대중 매체에서 진화라는 주제로 토론할 때, 사실은 그렇지가 않은데도 과학적 가설들이 너무나 자주 큰 권위와 신뢰를 얻고 있다. 전문가인 고생물학자가 하던 경고도 이제 사라지는 것 같다. 화석으로 보면 어류의 기원이 될 만한 중간형태가 하나도 보이지 않음으로써 문제가 되고 있다. 영국 BBC 방송의 '동물 탐험 시리즈(Zoo Quest)'에서 사람들에게 자주 언급되는 바로 그런 이야기(just-sostory)'에서 애튼버러(Dave Attenborough)는 다음과 같이 진술한다.
"기나긴 이 기간에 산호가 출현하고 그리고 산호초를 형성하기 시작하였고, 체절을 가진 동물이 곧 바다를 떠나 육지로 나갈 교두보를 확보하는 형태로 발전하였다. 중요한 변화가 또한 원시 어류 사이에서 일어났다. 물고기의 목구멍 양쪽 옆에는 여과 기능의 기원이 되는 아가미 틈새가 얇은 혈관으로 둘러싸이게 되어 아가미 역할을 하게 되었다. 이제 아가미 사이의 살기둥들이 뼈막대 형태로 딱딱해지고, 이 뼈들의 첫 쌍이 수천 년에 걸쳐 천천히 점진적으로 앞으로 향하면서 관절이 되었다. 그 주위로 근육들이 형성되어 막대 앞쪽 끝 부분이 위아래로 오르락내리락할 수 있게 되었다. 이렇게 해서 턱이 생겨난 것이다. 살을 덮는 피부에 골질의 비늘이 크고 날카롭게 자라고, 또 나중에는 이빨이 되었다. 이제 바다의
척추 동물은 더 이상 물을 여과하고 진흙을 걸러서 사는 미진화의 생물이 아니다. 이제 그들은 입으로 물 수 있게 되었다. 피부의 조직이 몸 아래 부위의 양쪽으로부터 생겨나서 물을 가르며 다니기에 편하게 하였는데, 이 부위가 실제로 지느러미가 되었다. 이제 어류는 수영할 수 있게 되었다. 그리고 처음으로 척추동물 사냥꾼들이 해수(海水)를 가로지르며 능숙하고 정확하게 스스로를 조정할 수 있게 된 것이다. "46)
그러나 우리는, 이 기나긴 시간 동안에 발생했을 것으로 제안된, 변화에 대한 화석 기록은 실제로 어디에도 존재하지 않는다. 그러나 TV 시청자들은 이 말의 문제점에 대하여 아무것도 듣지 못한다. 오히려 진화의 옹호자들은 한층 더 단언적이다. 에커(Ronald Ecker)는 기록하기를 "화석 기록이 불완전하고 커다란 간격이 많이 있다는 것은 확실하다. 그러나 이것이 결코 진화론을 믿을 수 없게 하는 것은 아니다."고 하였다. 그러한 예들은 어떤 확실한 증거 없이도 이 시대를 지배하고 있는 패러다임이 어떻게 지탱되고 있는지를 잘 표현해 주는 것이다. 49)
결론
화석은 생명의 진화를 호소하는 최후의 심판대이기는커녕 오히려 창조를 호소하는 최후의 심판대이다. 과학자들은 종종 새로운 개념은 소위 반증 검사를 해보아야 한다고 주장한다. 다시 말하면 개념이 잘못되었다는 것을 보여 줄 자료를 찾는 것이다. 진화가설이 잘못된 것임을 입증하는 한 가지 방법은 특히 주요 개체군(집단)들 사이의 화석이 지질 주상도 전반에 걸쳐서 연속성을 보이지 않는다는 것을 보여 주는 것이다. 만약 진화가 발생했다면 우리는 단순한 것에서부터 현재 생존하고 있는 모든 주요 형태의 생명체에 이르기까지 대부분의 화석이 연속적인 계열로 나타날 것을 기대할 수 있어야 할 것이다. 또한 생명의 돌연한 출현대신에 화석을 포함하는 하부 지층에서 서로 연관되어 있는 모든 주요 집단을 우리는 발견할 수 있어야 한다. 그러나 잘 알려진 대로 화석 기록은 중간형태를 갖고 있지 않다. 지질 주상도의 전반에 걸쳐서 동식물의 최고 분류단계인 문(門)을 넘어서 수백 종의 작은 단독 집단의 돌연한 출현으로까지 연장된다. 더구나 여기에 진화적인 변화에 시간이 거의 필요치 않았다는 상식을 벗어난 빠른 진화 속도의 문제점이 여기에 부가되어야 한다. 진화론에서는 캄브리아기 폭발과 같은 매우 복잡하고 불가능한 것처럼 보이는 주요 진화 발달이 단 몇 천만 년 사이에 일어난 것으로 가정하고 있다. 모든 자료를 종합적으로 볼 때, 일반적인 진화 모델은 본질적으로 거짓임을 말하고 있다.
■ 참고문헌
1) Wittgenstein L 1980. Culture and value. Winch P, translator: Wright GHv (with Nyman H), editor. Chicago: University of Chicago Press, p. 62e, Translation of: Vermischte Bemerkungen. 2) Lull RS. 1935. Fossils: what they tell us of plants and animals of the past, 2nd. New York: The University Society, p. 3.
3) 이 집단에 대한 기술은 제9장을 참고하고, 또 그 분포는 <그림 10.1>을 참고하라.
4) 일부 낙천적인 고생물학자는 캄브리아기 폭발 때에 100여 개 문의 등장을 제안한다. 다음을 참고하라. Lewin R 1988. A lopsided look at evolution, Science 241:291–293.
5) (a) Bowring SA, Grotzinger JP, Isachsen CE, Knoll AH, Pelechaty SM, Kolosov P. 1993, Calibrating rates of Early Cambrian evolution, Science 261:1293-1298. 182 414 (6) Nash M. 1995. When life exploded, Time 146(23):66-74.
6) Kerr RA 1995. Timing evolution's early bursts. Science 267:33, 34. 7) Stanley SM. 1981. The new evolutionary timetable: fossils, genes, and the origin of species, New York: Basic Books, p. 93.
8) Feduccia A 1995, Explosive evolution in Tertiary birds and mammals, Science 267:637, 638. 9) 제8장을 참고하라.
10) (a) Gould SJ. 1989. Wonderful life: the Burgess Shale and the nature of history. New York and London: WW Norton and Co., pp. 39-50 4. 412, (b) Briggs DEG. Fortey RA, Wills MA 1992. Morphological disparity in the Cambrian, Science 256:1670-1673: and later discussion by: (c) Foote M, Gould SJ, and Lee MSY. 1992, Cambrian and recent morphological disparity. Science 256:1816, 1817, with a response by Briggs, Fortey and Wills in Science 256:1817 1818.
11) Stewart WN, Rothwell GW. 1993. Paleobotany and the evolution of plants, 2nd ed. Cambridge and New York: Cambridge University Press, pp. 510, 511,
12) 무척추동물에서 종의 다양성은 퇴적암의 양과 분포지와 깊은 상관이 있다고 시사된다. 다음을 참고하라. (a)Raup DM, 1976, Species diversity in the Phanerozoic: an interpretation, Paleobiology 2:289–297: (b)Ey에서 보면 상부층으로 갈수록 노출된 퇴적암과 그 양이 많기 때문에, 보고된 종의 수도 많다고 생각할 수 있다. 그러나 그 기본형은 매우 적다. Raup DM, 1972, Taxonomic diversity during the Phanerozoic, Science 177:1065-1071.
13) 사례는 제4-8장을 참고하라.
14) Bernini F. 1991, Fossil Acarida, In: Simonetta AM, Conway Morris S, editors. The early evolution of Metazoa and the significance of problematic taxa. Cambridge and New York: Cambridge University Press, pp. 253-262
15) (a) Pennisi E. 1994, Static evolution: is pond scum the same now as billions of years ago? Science News 145:168, 169: (b) Schopf JW. 1968. Microflora of the Bitter Springs Formation, Late Precambrian, central Australia, Journal of Paleontology 42:651-688. 16) Stewart, Rothwell, p. 44 (111)
17) Knoll AH, 1990, Precambrian evolution of prokaryotes and protists, In: Briggs DEG, Crowther PR editors. Palaeobiology: a synthesis, Oxford and London: Blackwell Scientific Publications, pp. 9-16. 18) 제10장을 참고하라.
19) (a) Carroll RL 1988. Vertebrate paleontology and evolution, New York: W. H. Freeman and Co., p. 207. For an attempt at explaining the turtle evolution on embryological grounds but not paleontological data, see: (b) Petto AJ. 1983, The turtle: evolutionary dilemma or creationist shell game? Creation/Evolution 3(4):20-29. For an attempt at explaining the anatomy based on bones, see: (c) Lee MSY, 1993. The origin of the turtle body plan: bridging a famous morphological gap. Science 261:1716-1720,
20) Darwin C. 1859. The origin of species by means of natural selection, or the preservation of favoured races in the struggle for life, London: John Murray. In: Burrrow JW, editor, 1968 reprint.
ORIGINS
London and New York: Penguin Books, pp. 291, 292
21) Ibid., p. 309.
22) Raup DM, 1979. Conflicts between Darwin and paleontology. Field Museum of Natural History Bulletin 50:22-29.
23) Kitts DB, 1974, Paleontology and evolutionary theory. Evolution 28:458-472 24) Gould SJ. 1980. The panda's thumb: more reflections in natural history. New York and London: W.W. Norton and Co., p. 181.
25) 제8장을 참고하라.
26) 천해성 해양 동물의 경우, 모든 문(phyla)은 발견되었다고 코웬은 말한다. Cowen R 1995. History of lile.2nd ed. Boston, Oxford and London: Blackwell Scientific Publications, p. 97.
27) Simpson GG. 1967. The meaning of evolution: a study of the history of life and of its significance for man, Rev. ed. New Haven and London: Yale University Press, pp. 232, 233,
28) 진화론자, 창조론자 혹은 그 외의 사람들이 이 간격에 대하여 많이 기록하였다. 문제를 인식한 몇 사례는 다음 (a) Denton M. 1985, Evolution: a theory in crisis. London: Burnett Books: (b) Grassé P P. 1977. Evolution of living organisms: evidence for a new theory of transformation, Carlson BM, Castro R translators, New York, San Francisco, and London: Academic Press. Translation of: L'É volution du Vivant: (c) Hitching F. 1982. The neck of the giraffe: where Darwin went wrong. New Haven and New York: Ticknor and Fields: (d) Hoffman A. 1989. Arguments on evolution: a paleontologist's perspective. New York and Oxford: Oxford University Press: (e) Johnson PE, 1993. Darwin on trial, 2nd ed. Downers Grove, III: InterVarsity Press: (1) Løtrup S. 1987. Darwinism: the refutation of a myth, London, New York and Sydney: Croom Helm: (g) Pitman M. 1984, Adam and evolution, London, Melbourne and Sydney: Rider and Co.
29) Schram FR 1991. Cladistic analysis of metazoan phyla and the placement of fossil problematica In: Simonetta and Conway Morris, p. 35-46 (14).
30) Bold HC, Alexopoulos CJ, Delevoryas T. 1987. Morphology of plants and fungi, 5th ed. New York and Cambridge: Harper and Row, p. 823.
31) (a) Axelrod DI. 1960. The evolution of flowering plants, In: Tax S, editor. The evolution of life: its origin, history and future. Evolution after Darwin: The University of Chicago centennial, Vol. 1, Chicago: University of Chicago Press, pp. 227-305: (b) Bold HC, 1973. Morphology of plants, 3rd ed. New York and London: Harper and Row, p. 601 (원본의 4판과 5판은 2명의 공동저자로 되어 있고 '지독한'이라는 용어는 더 이상 사용되지 않았다. 그러나, 책에서 그의 생각은 여전하다. (c) Knoll AH. Rothwell GW. 1981, Paleobotany: perspectives in 1980. Paleobiology 7(1):7-35. 32) Wootton RJ. 1990. Flight: arthropods. In: Briggs and Crowther, pp. 72-75 (17)
33) Gibson LJ. Are the links still missing? Unpublished paper distributed by the Geoscience
Research Institute, Loma Linda University, Loma Linda, California 34) There has been a dispute over the authenticity of the Archaeopteryx fossils, but they appear authentic. See: (a) Charig AJ, Greenaway F. Milner AC, Walker CA, Whybrow PJ. 1986. Archaeopteryx is not a forgery. Science 232:622-626: (b) Clausen VE, 1986. Recent debate over Archaeopteryx Origins 13:48-55,
35) (a) Wheeler TJ. 1993. Were there birds before Archaeopteryx? Creation/Evolution 13(2):25-35; (b) Zimmer C. 1992, Ruffled feathers. Discover (May), pp. 44-54. 36) 논쟁의 발단에 대하여 제9장을 참고하라.
37) Simpson GG, 1953. The major features of evolution, New York and London: Columbia University Press, p. 263.
38) Raup 1979 (22)
39) Milner R 1990. Horse, evolution of. The encyclopedia of evolution, New York: Facts on File, p. 222 40) MacFadden BJ. 1992. Fossil horses: systematics, paleobiology, and evolution of the family equidae, Cambridge and New York Cambridge University Press, p. 330.
41) Kemp TS. 1982, Mammal-like reptiles and the origin of mammals. London and New York:
제11장 - 진화에 대하여 화석이 말하는 것 | 209
Academic Press, p. 296.
42) Carroll, p. 398 (19a).
43) Patterson C. 1978. Evolution, London: British Museum (Natural History), and New York: Cornel University Press, p. 133. Patterson lists this explanation but does not especially defend it
44) Ibid
45) Kerr RA 1991. Old bones aren't so bad after all, Science 252:32, 33.
46) Paul CRC. 1990. Completeness of the fossil record. In: Briggs and Crowther, pp. 298-303 ( 17).
47) (a) Denton, p. 190 (28a). Denton (b) Romer AS. 1966. Vertebrate paleontology. 3rd ed. Chicago and London: University of Chicago Press, pp. 347-396. 48) Attenborough D. 1979. Life on Earth: a natural history. London: William Collins Sons and the British Broadcasting Corporation, p. 112
49) Ecker RL 1990. The dictionary of science and creationism, Buffalo: Prometheus Books, p. 94.