Memory
As shown in Figure 03a, the ability to modify our behaviour in response to life's experiences is shared by all animals including the bacteria E. coli. Such feat requires the brain's willingness to learn. Learning results in the formation of memories and in humans this process reaches its most sophisticated form, allowing us creatively to associate different reflections on the past, to generate new ideas, and most importantly to acquire language as a medium of expression and communication. Memory requires the brain to be physically altered by experience and it is this remarkable property that makes thought, consciousness, and language possible. The basic mechanism of memory formation is highly conservative over
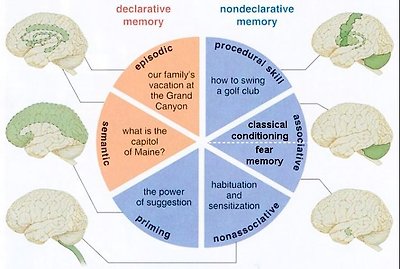
Figure 24a Memory Classification
billion years of biological evolution. The difference in humans is that we have a lot more of the stuffs. There are about 100 trillion synaptic connections in our brain.
There are many ways to classify the memory. The concept of explicit and implicit memory refers to whether or not the recollection is produced consciously and intentionally. While the scheme of declarative and nondeclarative memory depend on the retrieval that can be declared verbally or not. Associative memory is triggered by clues; nonassociative memory can be habitual or sensitive. There are also short term and long term memory. One of the classification schemes is shown in Figure 24a. Table 07 is an attempt to put them all together. In the table, the declarative, and the procedural memory are explicit with the rest of nondeclarative memories being implicit. Only the working memory belongs to the category of short term memory fading away in hours, while the others are long term, and available for retrieval in years. Figure 24b shows the components, locations, and pathways for many types of memory.
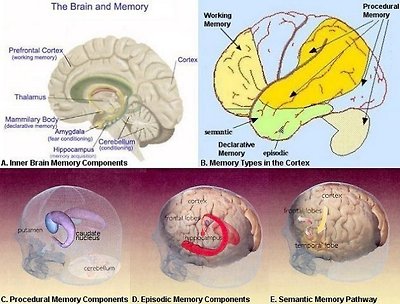
Figure 24b Types of Memory
| Type | Location(s) | Function | Example(s) |
|---|---|---|---|
| Working Memory | |||
| Phonological Loop | Left hemisphere | Rehearsing verbal information to keep it in the short-term memory | String of numerals and alphabets such as telephone numbers |
| Visual-spatial Scratch Pad | Visual Cortex | Controlling visual imagery | Scanning text |
| Central Executive | Frontal lobe | Controlling awareness of the information in working memory | Constructing sentence, comprehending speech |
| Non-declarative Memory | |||
| Procedural Memory | Cerebellum, temporal lobes | Managing "how to" | Riding a bicycle, kungfu exercise |
| Classical Conditioning | Cerebellum | Forming habitual behaviour | Coffee break, afternoon tea |
| Fear Memory | Amygdala | Emotional conditioning | Phobias, flashbacks |
| Nonassociative Memory | Spinal cord | Habituation and Sensitization | Decreased or increased responsiveness to stimulus |
| Remote Memory (Priming) | Scattered around the cortex | Foundation for new memories | Childhood memory |
| Declarative Memory | |||
| Episodic Memory | Cortex | Remembering past experience | Some enchanted evening |
| Semantic Memory | Frontal lobe, temporal lobe | Registering facts | Meanings of words and symbols |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 07 Types of Memory
•Working Memory -Most of our memories are fleeting because few of the many experiences we have in the course of an average day are remembered for very long, nor do they need to be. Transient memories are absolutely essential to the process of understanding the meaning of events as they occur in the present. This type of very short-term memory for things being experienced now is known as working memory; it allows you to comprehend what you are reading or to figure out the meaning of what has just been said to your in a conversation. Working memory can be thought of as a low capacity information reservoir that is always full, sensations flowing into it continuously at about the same rate that they are forgotten. Some of the information held in short-term storage may be important enough to be remembered for a long time and must therefore be transferred to a more stable form of storage, which is represented by far more robust alterations in the brain's chemical and physical make-up in the form of synaptic connections. It is not necessarily for an important experience to trigger the formation of long-term memories, other factors such emotion, practice, and rehearsal also facilitate the transformation. Experiments show that in all cases the most important underlying distinction between the short- and long-term memory formation is that the latter requires a dialogue between synapses and genes and the former does not.
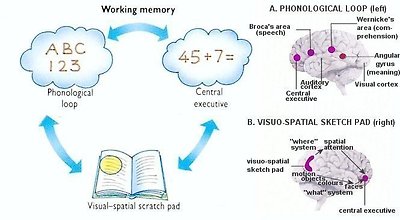
Figure 25 Working Memory
The working memory itself is located in the prefrontal cortex. As experimental techniques became refined, it has become clear that there is no rigid dividing line between a memory and a thought. A model of working memory has been developed to combine perceptions, memories and concepts together, and consists of three parts:
◦Phonological loop - Memory in this area (see Figure 25) enables us to remember sequences of approximately seven digits, letter, or words. The language areas of the brain are mainly in the left hemisphere, around and above the ear. The language loop start with hearing words in the auditory cortex and/or reading words in the visual cortex. Perception of language results from the convergence of auditory and visual information in Wernicke's area. Expression of language is controlled by Broca's area; while the angular gyrus is concerned with meaning.
◦Visual-spatial scratch pad - It is like a sort of inner eye, which receives and codes data into visual or spatial images. For example, it comes into play when we need to remember where we were on a page when we start reading a book again. Functional imaging suggests that this complex structure represents the "what" and "where" in short-term memory (see Figure 25).
◦Central executive - This most important yet least well understood component of the working memory model, is postulated to be responsible for the selection, initiation, and termination of processing routines (e.g., encoding, storing, retrieving). It is believed that this component coordinates information from a number of sources, directs the ability to focus and switch attention, organizes incoming material and the retrieval of old memories and combines information arriving via the other two temporary storage systems. It performs various tasks such as reasoning or doing mental arithmetic - rather like the RAM (Radom Access Memory) of a computer.
•Nondeclarative memory - Nondeclarative memory includes skill learning, implicit learning, priming, simple classical conditioning, and habituation. These forms of learning are similar in that it is experience which changes the neural makeup, and the conscious access to past episodes is not essential for the formation of these memories. Implicit memory is not flexible and does not allow for the recombination of learned information. Nondeclarative memory does not require the hippocampus or related structures. Instead, the implicit learning of skills and habits depends on the neostriatum (basal ganglia and its connections to the frontal lobes). The conditioning simple skeletal muscle reactions depends on the cerebellum. The amygdala is
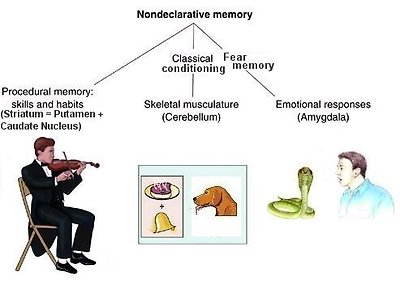
Figure 26 Nondeclarative Memory
essential for emotional conditioning. Nondeclarative memory can be classified to five main groups:
◦Procedural memory - It is the repository of such skills as handwriting or driving. These skills are essential part of our memory store, but it is difficult to describe the "know-how" in words. In this sense the memory is said to be implicit or non-declarative (Figure 26); you just cannot explain how to ride a bicycle. The skills may be difficult to acquire, but once learnt they are never forgotten, even without occasional practice. Thus it seems that the know-ledge or information required for the execution of very complex motor routines or procedures is somehow laid down in a robust permanent memory store. The parts of the brain involved in the acquisition of complex motor skills are the cerebellum and putamen (see Figure 24). Deeply ingrained habits are stored in the caudate nucleus.
◦Classical conditioning - Along with motor skills, conditioning is part of non-declarative memory. The desire for food at a particular time of day - regardless of whether hungry or not - is one example of such conditioning. A classical example is to associate the ring of a bell to food when feeding a dog. After repeating the training many times, the dog shows salivation at the ring of the bell even without food (see Figure 26).
◦Fear memory - Recent study in delivering shocks to mice suggests that fear memory does not occur immediately after a painful event; rather, it takes time for the memory to become part of our consciousness. The initial event activates NMDA receptors - molecules on cells that receive messages and then produce specific physiological effect in the cell - which are normally quiet but triggered when the brain receives a shock. Over time, the receptors leave their imprint on brain cells. A phobia is an excessive or unreasonable fear of an object, place or situation. Examples include fears of specific things such as insect, snake, mouse, and flying. It seems that people can learn to suppress a fright reaction by repeatedly confronting, in a safe manner, the fear-triggering memory or stimulus. It is found that for specific phobias, up to 90% of people can be cured through such exposure therapy.
◦Nonassociative memory - Nonassociative memory includes two forms of learning called habituation and sensitization. Habituation is defined as a decreased in response to a repeated stimulus such as a certain odor. On the other hand, sensitization is an increased responsiveness such as more sensitive in touching a cut in the skin. Nonassociative learning involves reflex pathways in the spinal cord and elsewhere.
◦Remote memory - The memory of events that occurred in the distant past is referred to as remote memory. The underlying anatomy of remote memory is poorly understood, in part because testing this type of memory must be personalized to a patient뭩 autobiographical past. What is known is that, like semantic memory, remote memory eventually becomes independent of the hippocampus. One memory model shows a linear representation of how experience is processed as memory: Stimulus Sensory Registration Attention Short Term Memory Consolidation - Retrieval Long Term Memory Remote Memory. At the stage of sensory registration, there is a matching/assigning of the pattern to a meaning. Short-term memory is temporary and is limited in space. If short-term memory is not repeated, the information is lost fairly quickly. Long term memory is consolidated and stored throughout the nervous system. Remote memories represent the foundation memories upon which more recent memories are built. Since early acquired information is the foundation for new memories and may be linked to many more new memories, such memory is less subject to change and/or loss. Similar to the short-term memory, the remote memories are not usually affected by aging.
•Declarative memory - Declarative memory covers the memory of facts such as events and names, which do not need to be repeated for them to sink in. Those experiences destined to be laid down as long-term memories are shunted down to the hippocampus where they are held in storage for 2 - 3 years. During this time the hippocampus replays the experiences back up to the cortex, and each rehearsal etches it deeper into the cortex. Eventually the memories are so firmly established in the cortex that the hippocampus is no longer needed for their retrieval. Much of the hippocampal replay is thought to happen during sleep. Dreams consist partly of a rerun of things that have happened during the day, fired up to the cortex by the hippocampus. The visual areas generate rerun of daily sightings (Figure27).
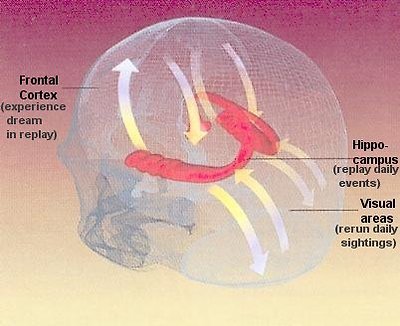
Figure 27 Long-term Memory
Semantic, and episodic memory are the subclasses of declarative memory:
■Episodic memory - It is about an event in one's life and everything about it, including emotional reactions. Remembering an episode, e.g., the attack on Pearl Harbour (Figure 28), is to create a memory for a unique event that only happened once and there is no opportunity for learning the event by rehearsal. Episodic memories are not very reliable, they are highly personal, selective, idiosyncratic and varying over time, but they may also be richly complex and movie-like in character. They constitute the stories we tell ourselves about our past, they are the things we would write about in our autobiography. Episodic memories can be recalled deliberately or are triggered by evocative sensory stimuli - particularly by the sense of smell. Episodic memory involves the use of the hippocampus for forming memories and the cortex for storage (see diagram D, Figure 24).
Figure 28 Declarative Memory
◦Semantic memory - Semantic memory is the knowledge of facts - numbers, addresses, language and concepts - which the brain files in categories and which seems to involve the left temporal lobe. Retrieval is then carried out by the frontal lobes (see diagram E, Figure 24). We assume all of the facts that constitute our knowledge of things must be stored in an organized fashion to be useful. Though this has not been demonstrated, it seems likely that the brain stores our semantic memories as modules that have some logical links to one another; they are grouped by category for instance. On retrieval, the brain knows where to find the memory according to the address of that particular category. Semantic memory is essential to the understanding of how things work and thus to an under-standing of the world we live in. It is a body of knowledge that helps us to regulate our behaviour according to and dependent on reliable factual memories. Navigational skills, for example, depend on our ability to deploy a complex store of semantic memory, including detailed spatial memories and representations of the world.
Memory is created by association between a group of neurons such that when one fires, they all fire, producing a specific pattern. Thought, sensory perceptions, ideas, and hallucinations - any brain function is made up of this same thing. For example, a group of neighbouring neurons firing together in the auditory cortex would bring about the experience of a certain note of music. A memory is a pattern like these. The only difference is that it remains encoded in the brain after the stimulation that originally gave rise it has ceased. Memories form when a pattern is repeated frequently, or in circumstances that encourage it to be encoded. This is because each time a group of neurons fires together the tendency to do so again is increased. Once the neighbour has been triggered to fire a chemical change takes place on its surface which leaves it more sensitive to stimulation from that same neighbour. This process is called long-term potentiation (LTP). If the neighbour cell is not stimulated again it will stay in this state of readiness for hours, maybe days. If the first cell fires again during this period, the neighbour may respond even if the firing rate of cell number one relatively slow. A second firing makes it even more receptive and so on. Eventually, repeated synchronous firing binds neurons together so that the slightest activity in one will trigger all those that have become associated with it to fire, too. A memory has been formed.
The giant sea slug called Aplysia californica¶ is often chosen for the studying of memory. Its brain has about 20000 neurons, some of which are large enough to be visible to the naked eye. Aplysia can learn and most importantly it is found that the mechanisms and principles involved in its formation of short- and long-term memories are conserved throughout the animal kingdom, including in humans. Aplysia exhibits a behaviour of protective reflex in which the sea slug withdraws its gill into the safety of the mantel cavity in response to a mild touch stimulus to another part of the body called the siphon (Figure 29a). If the stimulus is repeated a number of times, the gill withdrawal reflex becomes weaker until finally the animal ignores the touch stimulus. The waning of sensitivity to repeated stimulation is known as habituation and is a very simple form of learning found in all animals, including humans. Another type of learning is sensitization, when we are exposed to an unexpected or strongly unpleasant stimulus. Generally the sensitizing effect of
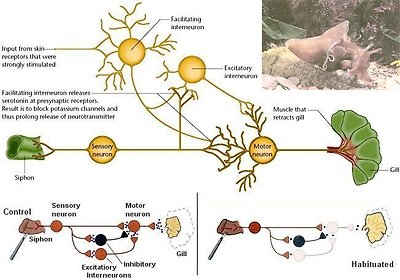
Figure 29a Memory in Aplysia
a single alarming stimulus is short-lived, lasting perhaps for just a few minutes. But if the alarming stimulus is repeated a number of times our senses may be heightened for days and now such sensitization becomes a form of long-term memory.
It turns out that short term changes involves only modification of pre existing proteins and alterations of pre existing connections. The short term process does not involve ongoing macromolecule synthesis. The effect wears off with time or repeated applications with no untoward happening. On the other hand, long term process involves a structural change which is not seen in the short term. In long term processes, there is a growth in new synaptic connections by sensory neurons onto follower cells. The growth of new synaptic connections is activated by the gene expression resulting in new protein synthesis.
At the macromolecule level, it is known that the neurotransmitter involved in the processes is the serotonin. A puff of serotonin alone can substitute for the siphon shock. It is shown further that the serotonin triggers the release of the second chemical messenger called cyclic-AMP. It activates an important type of enzyme called a kinase, which modifies the properties of particular target proteins by adding a phosphate molecule to them; the term for this is protein phosphorylation. The target for this modification in the sensory neuron is a potassium channel protein. It is mentioned earlier that a potassium channel is important in the downward phase of the action potential. The net result of phosphorylation is a prolongation of the action potential in the sensory neuron and so more neurotransmitter is released by the sensory neuron. Thus the sensory neuron's synapse with the gill motor neuron is strengthened. In short-term memory, special enzymes quickly remove phosphates from the proteins and return them to their original state, restoring the synaptic strength to its lower pre-sensitized level. However, following repeated serotonin delivery, the level of cAMP-activated kinase is much higher and this allows the crucial step in the formation of long-term memory to occur. This crucial step is the transport from the synapse to the cell body of kinase molecules that have been activated by c-AMP. Once in the cell body the activated kinases enter the nucleus and start to regulate the expression of particular genes. In Aplysia, proteins that result from this process of gene activation are transported back to the synapse where they are used to maintain the strength of synapses already affected by local effects of c-AMP and to grow new synaptic connections. So in Aplysia (as in other animals) the conversion of a short-term into a long-term memory involves the reinforcement of the short-term changes in synaptic strength and the growth of new synapses, both of which require the synthesis of new proteins.
As mentioned earlier, repeated activation yields a lasting increase in the efficiency of synaptic transmission -- a process called long-term potentiation (LTP) -- which is thought to underlie memory formation. LTP depends both on enhanced insertion of receptors for the neurotransmitter glutamate at spines (the sites of synapses) and on spine growth. Figure 29b shows the molecular basis of LTP as described in a 2008 neuroscience article. Essentially, it reports that the process is driven by the myosin V proteins, which shuttling receptors and membranes to make synaptic junctions better detectors of incoming signals.
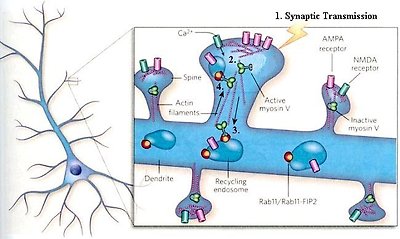
Figure 29b LTP, Molecular Basis of
A brief description for each step of the process is outlined in the following (also see
Figure 29b):
1.Neurons receive synaptic transmission from other neurons (not shown) at dendritic spines.
2.Influx of Ca2+ through NMDA receptors activates the folded (inactive) form of the myosin V motor protein.
3.The active myosin V protein (in extended form) then moves to the dendrite shaft and binds to Rab11/Rab11-FIP2 on recycling endosome (reused membraneous sac) containing AMAP receptors.
4.Finally, it transports the cargo into and along spines via actin filaments (microtubules) to mediate insertion of AMPA receptors at the cell surface, as well as spine growth through membrane insertion.
It is reported in 2007 that the seat of memory has been pinpointed in mouse. By monitoring 260 neurons in the hippocampus (Figure 29c), researchers have discovered that different experience is recorded in different area called "clique", which can be categorized from very general to very specific. Furthermore, such brain activities can be translated into binary codes (Figure 29d). Supposedly, we can read the mind from such codes and tell what it is thinking by the process of
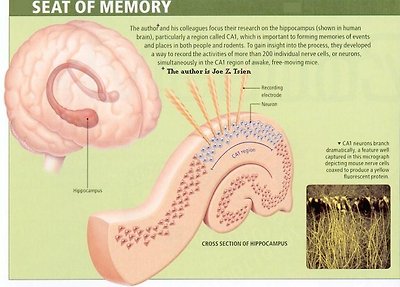
Figure 29c Seat of Memory
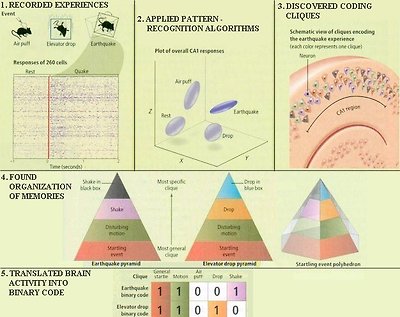
Figure 29d Memory Code
backward translation. The followings are steps to uncover the memory code:
1.Recorded Experiences - The mice are exposed to three startling experiences - a puff of air on the back (to mimic an owl attack from the sky), a fall in a container (the "elevator" drop), and shaking in a cage (the "earthquake") - while a recorded plotted firing from a large set of CA1 neurons. Each row in the plot captures firing of a single cell over time.
2.Patterns of Mental Activities - The points in the 520 dimensional phase space (corresponding to the activities of 260 neurons before and after an event) are projected into a 3 dimensional phase space. Different mental activity falls into different area in such plot starting from "rest". Temporal analysis revealed that the activity patterns associated with those startling experiences recurred spontaneously at intervals ranging from seconds to minutes after the actual event, but with smaller amplitudes than the original response. Such patterns provide evidence that the information traveling through the hippocampal system was inscribed into the brain's memory circuits. The replay corresponds to a recollection of the experience after the fact.
3.Coding Cliques - It is discovered that neuron ensembles active during an event contain subsets -termed neural cliques. The cells in a clique all show very similar firing patterns and are not part of the other cliques.
4.Organization of Memories - Further analyses showed that each clique encodes a different aspect of an experience, ranging from the general to the specific. It can be visualized as a hierarchical organization with the most general clique at bottom, and the very specific on top. Any given pyramid can be a component of a more general polyhedron representing all events of a given category, such as "all startling events".
5.Translated into Binary Code - The clique activity is represented as a string of binary code with 1 as being active and 0 signifies inactivity. Thus the earthquake binary code is 11001 corresponding to: "starting event", "disturbing motion", "air puff", "drop", and "shake". While the elevator drop binary code is 11010 for the same sequence.
